| Головна |
| «« | ЗМІСТ | »» |
|---|
В основі сучасних уявлень про структуру і функції мозку лежить нейронна теорія. Незважаючи на те що нервові клітини, з яких побудований мозок, складають єдину організовану мережу, структура ЦНС настільки складна, що для аналізу її функціональних властивостей доцільно поділ всього мозку на певні відділи відповідно до їх розвитком, функцією, нейронної організацією і макроструктурою. Такий поділ умовно, так як нейрони з віддалених відділів мозку бувають пов'язані між собою тісніше, ніж територіально ближче розташовані. Таке умовне розділення ЦНС зручно як для клінічних цілей, так і для нейрофизиологічного аналізу.
Спинний мозок філогенетично є найдавніше освіту ЦНС. Безліч і різноманітність рефлекторних процесів (рухових і вегетативних), що здійснюються спинним мозком, зумовили його складну структуру, різноманітні форми нейронів і міжнейронних зв'язків. Спинний мозок укладений в хребет і має сегментарну будову. Від кожного сегмента відх
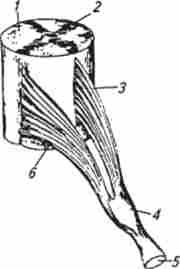
дять дві пари вентральних і дорсальних корінців, які з'єднуються і утворюють периферичні спинальні нерви.
аферентні волокна вступають в спинний мозок через задні корінці, а все еферентні волокна виходять з нього через передні корінці відповідно до закону Белла-Ма-Жанден (рис. 13.10). З його відкриттям вперше був установлений розподіл нервових воло-
Мал. 13.10. Передні і задні спинномозкові корінці та спинальний ганглій:
/ - біла речовина; 2 сіра речовина; J - передній корінець; 4 спинальний ганглій; 5-змішаний нерв; б-задній корінець кін на аферентні і еферентні. Роздратування центрального відрізка будь-якого заднього спинномозкового корінця викликає сильну больову реакцію, а роздратування його периферичного відділу не супроводжується відповідною рухової реакцією. Роздратування ж центральної ділянки переднього корінця, навпаки, не супроводжується реакцією, а роздратування його периферичного відрізка завжди викликає скорочення певних м'язів. Перерезка задніх корінців веде до втрати чутливості, а перерезка передніх корінців викликає параліч мускулатури.
Надалі було показано, що в передніх корінцях проходять крім моторних нейронів скелетної мускулатури інші еферентні нервові волокна - судинні, секреторні і йдуть до очних м'язів, що не суперечить закону бел ла-Мажанді.
Кожна пара спинномозкових корінців іннервує певні ділянку шкіри тіла і скелетні м'язи. Сегменти спинного мозку і їх корінці позначаються латинськими літерами, які вказують на міжхребцеві отвори, через які виходять корінці. Шийні хребці позначають буквою С (цервікальні), грудні - буквами D і Th (спинні, торакальні), поперекові - буквою L (люмбальні), крижові - буквою S (сакральні).
На поперечному перерізі спинного мозку центрально розташоване сіра речовина має своєрідну форму «метелики». У ньому розрізняють вентральні і спинні роги; останні включають в себе крайову зону, желатинозную субстанцію і підстава дорсального рогу. З боків сірої речовини знаходяться латеральні роги, які добре розвинені в грудному відділі мозку. Між Дорсально і вентральними рогами лежить проміжна зона.
Принцип класифікації нейронів спинного мозку запропонував Рекседа. Сіра речовина він розділив на 10 пластин (рис. 13.11).
1 ... 4-Я пластини утворюють «головку дорсального рогу» і є першою сенсорної зоною сірої речовини. З цієї області починається кілька трактів (спинно-таламичний та ін.), Що передають екстерорецептівние чутливість до вищих центрам.
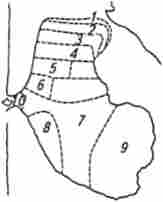
5-я і 6-я пластини утворюють «шийку дорсального рогу». У цій області локалізуються численні типи вставних нейронів, які отримують входи від волокон заднього корінця і низхідних шляхів (кортикоспинального і руброспінальний тракту).
Мал. 13.11. Пошарова топографія сірої речовини спинного мозку по Рекседа. цифрами позначені шари нервових клітин
У 7-й і 8-й пластинах локалізуються пропріоспінальние вставні нейрони, що дають початок довгим аксонам. Тут закінчуються аферентні волокна від пропріорецепторов, волокна вестібулоспінальних і ретикулоспинального трактів, аксони пропріоспінальних нейронів.
Пластина 9 є первинної моторної областю сірої речовини і складається з мотонейронів спинного мозку, об'єднаних у функціональні групи. Це кінцевий спільний шлях рефлексів спинного мозку.
Пластина 10 оточує спинномозковий канал і містить поряд з нейронами значне число гліальних клітин і комісуральних волокон.
Рефлекторна діяльність спинного мозку. У спинному мозку замикається дуже багато рефлекторних дуг, за допомогою яких регулюються як соматичні, так і вегетативні функції організму. На його різних рівнях розташовані центри, які беруть участь в регуляції рухів всіх м'язів голови, шиї, тулуба та кінцівок. На рівні 3 ... 5-го шийних хребців знаходиться центр скорочення діафрагми, а в крижовому відділі -? центри дефекації і сечостатевих рефлексів. Так як від спинного мозку відходять і вегетативні нерви, то він грає велику роль в регуляції судинного тонусу, тканинного обміну, розширення зіниць, відділення поту.
Рухові рефлекси спинного мозку здійснюються через альфа-мотонейрони передніх рогів. Дуги цих рефлексів можуть бути моносинаптічеськие і полісинаптичні.
Таким чином, рефлекторна діяльність спинного мозку - це сукупність елементарних рухових рефлексів і рефлексів підтримки пози. Ізольованих рефлексів тольк' спинного мозку не існує. Функціонування і взаємодія всіх рефлексів залежать від регулюючих впливів розташованих вище центрів.
Провідникова функція спинного мозку. Зв'язок спинного мозку з встановленими вище відділами ЦНС (стволом мозку, мозочком і великими півкулями) здійснюється за допомогою висхідних і низхідних провідних шляхів.
Висхідні провідні шляхи спинного мозку складаються з пучків нервових волокон, які проводять імпульси від рецепторів вгору по спинному мозку до верхніх відділів ЦНС. Волокна висхідних шляхів - це або аксони клітин спинального ганглія, або аксони тих нервових клітин сірої речовини спинного мозку, у яких закінчуються розгалуження волокон аферентних нейронів. Провідні шляхи являють собою пучки волокон, що утворюють білу речовину стовпів спинного мозку. При цьому під передніми стовпами спинного мозку розуміють масу білого речовини між передній щілиною спинного мозку і передніми корінцями; бічні стовпи - це біла речовина, обмежене передніми і задніми рогами сірої речовини спинного мозку і відповідними корінцями; задні стовпи - біла речовина між задніми рогами сірої речовини і задніми корінцями. До шляхах відносять такі: пучки Голля і Бурдаха; пучки Флексига і Говерса;
спинно-таламичні тракти (латеральний і вентральний). Величезна маса волокон, що йдуть від м'язових рецепторів, проходить в медіальній частині задніх корінців і, вступаючи в спинний мозок, утворює пучок Голля і пучок Бурдаха, які складають задні стовпи спинного мозку (рис. 13.12).
Пучки Голля і Бурдаха є відростками клітин спінальних гангліїв, висхідними в складі задніх стовпів до довгастого мозку. У довгастому мозку волокна пучків закінчуються у однойменних ядер Голля і Бурдаха. Від клітин цих ядер відходять нові волокна до верхніх центрам. Крім волокон, що йдуть від м'язових рецепторів, в пучках Голля і Бурдаха сходить
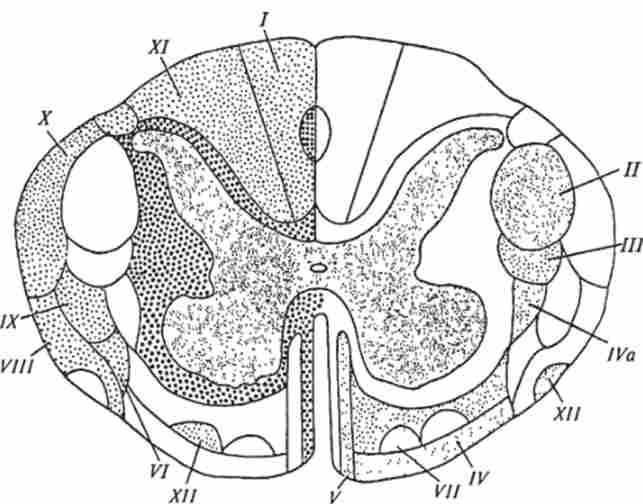
Мал. 13.12. Провідні шляхи спинного мозку. відзначено иева точками висхідні, праворуч - спадні шляху; кружками - міжсегментні шляху; висхідні шляху:
/ -пучок Гслля; XI- пучок Бурдаха; А'-дорсальний спинно-мозочковою тракт Флексига; VIII- вентральний спинно-мозочковою тракт Говерса і VI-латеральний і вентральний спинно-таламічна тракт; XII - спинно-тсктальний тракт. спадні шляху (праворуч відзначені точками). // і К-бічний і передній пірамідний тракт; /// - руброспінальний тракт; IV- всстібулоспінальний тракт; VII- Олівоспінальний тракт також частина волокон від рецепторів шкіри, що сприймають роздратування при тиску і дотику, - тактильних рецепторів, а також від рецепторів вібраційної чутливості. Невелика частина цих волокон доходить до ядер Голля і Бурдаха, а решта закінчуються, віддавши ряд колатералей, в сірій речовині спинного мозку. Волокна заднього стовпа не перехрещуються в спинному мозку.
Спинний спинно-мозочковою тракт, або пучок Флексига, розташовується в задненаружной частини бічного стовпа спинного мозку. Волокна даного тракту є аксонами клітин тієї ж, але почасти й протилежного боку спинного мозку. Ці клітини лежать в сірій речовині поблизу підстави заднього рогу, утворюючи так званий стовп Кларка. Волокна дорсального спинно-мозочкового тракту закінчуються у клітин кори мозочка. Вентральний спинно-мозочковою тракт, або пучок Го- версія, проходить в передненаружной частини бічного стовпа спинного мозку, під пучком Флексига. Волокна цього тракту є відростками нервових клітин задніх рогів спинного мозку. Обидва спинно-мозочкових тракту складаються, отже, з відростків клітин, що лежать в самому сірій речовині спинного мозку. Всі волокна пучків Флексига і Говерса проводять до мозочка імпульси, що виникають в клітинах спинного мозку під впливом роздратування, переданого в центри від м'язових рецепторів. Пучок Флексига в основному не перехрещується в спинному мозку, а пучок Говерса перехрещується.
Латеральний спинно-таламічна тракт розташовується в задньому розі того ж сегмента, в який вступають безмякотние волокна больової, температурної і частково тактильної чутливості. Закінчується він у ядрах зорових горбів, перехрещується в спинному мозку, головним чином на рівні вступу аферентних волокон.
Вентральний спинно-таламічна тракт може розташовуватися в задньому розі будь-якого сегмента мозку, до якого доходять волокна відчуття дотику, а закінчується в ядрах зорових горбів; перехрещується в спинному мозку.
Волокна клітин задніх рогів, що переходять на протилежну сторону, утворюють спинно-тектальний тракт і закінчуються в ядрах четверохолмия.
Таким чином, волокна аферентного нейрона з клітинним тілом, лежать в спинальному ганглії, що не сходять до кори великих півкуль.
Спадні провідні шляхи спинного мозку включають в себе:
кортікоспінальних, або пірамідні, шляхи;
руброспінальний тракт (пучок Монакова, або екстрапіра- мідно);
вестібулоспінальний тракт.
Кортікоспінальних, або пірамідні, шляхи є відростками великих пірамідних клітин (клітин Беца) рухової зони кори великих півкуль. Велика частина волокон пірамідного шляху переходить на протилежну сторону в нижній частині довгастого мозку (перехрещення пірамід) і утворює латеральний кортикоспінального тракт (контралатеральний пірамідний тракт), розташований всередині від пучка Флексига в бічному стовпі спинного мозку. Менша частина волокон пірамідного шляху утворює передній кортикоспінального шлях (прямий, або непере- хрещений, пірамідний шлях), волокна якого утворюють вузький пучок, що лежить в передньому стовпі білої речовини спинного мозку. Однак і ці волокна переходять на протилежну сторону на рівні того сегмента спинного мозку, в якому закінчуються дані волокна.
Таким чином, клітини рухової зони кори великих півкуль пов'язані волокнами кортікоспінальних трактів тільки з протилежного боку спинного мозку; закінчуються волокна кортикоспинального тракту у рухових клітин передніх рогів, які віддають рухові волокна до скелетних м'язів. Пірамідний шлях розвинений тим сильніше, чим вище організована кора мозку. Максимального розвитку цей шлях досягає у людини. Мієлінізація пірамідного шляху закінчується лише до кінця другого року життя.
Руброспінальний тракт (Монакова) утворюється волокнами, які є відростками клітин червоного ядра середнього мозку. Ці волокна еше в середньому мозку повністю переходять на протилежну сторону (так званий перехрестя Фореля), а в спинному мозку проходять попереду від латерального кортикоспинального шляху в бічному стовпі спинного мозку. Руброспінальний тракт проводить імпульси від мозочка, ядра вестибулярного нерва і смугастого тіла до нейронів спинного мозку.
Вестібулоспінальних тракти утворені відростками клітин, що лежать в покришці середнього мозку, в оливі і в латеральному ядрі вестибулярного нерва. Волокна цих трактів закінчуються в вентральних рогах спинного мозку. За ним йдуть імпульси від вестибулярного апарату і мозочка до мотонейронам, що регулюють тонус м'язів, рівновагу і узгодженість рухів.