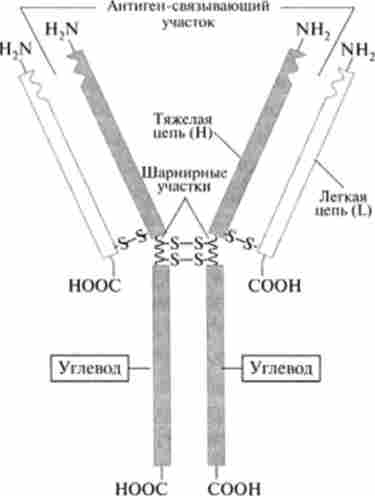
Антитіла є імуноглобулінами, що складаються з важких і легких поліпептидних ланцюгів. Зазвичай імуноглобулін позначають як Ig,
Мал. 30.8. Структура молекули імуноглобуліну G
а потім буквою вказують його приналежність до певного класу. IgG, на прикладі якого розглянемо структуру антитіл, має Y-подібну форму. Він складається з двох легких і двох важких поліпептидних ланцюгів, з'єднаних дисульфідними зв'язками (рис. 30.8).
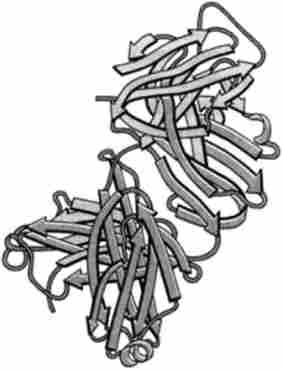
Важкі (Н) ланцюги складаються з 440, а легкі (L) - з 220 амінокислотних залишків. Кожна ланцюг має константні і варіабельні області. Варіабельні області локалізовані на yV-коніах обох ланцюгів, і саме вони утворюють ділянку зв'язування антигену. Різні варіанти амінокислотних послідовностей в ділянці зв'язування антигену обумовлюють структурну різноманітність сайтів взаємодії з антигенами. Не всі варіабельні області легких і важких ланцюгів контактують з молекулою антигену, а лише невеликі ділянки, що складаються з 20-30 амінокислотних залишків, так звані гіперваріабельні ділянки. Кожна ланцюг складається з повторюваних послідовностей амінокислот, т. Е. Доменів. Легка ланцюг складається з одного вариабельного (V,) і одного константного (CL) Доменів. І легкі, і важкі ланцюги містять по одному варіабельний домен. Що стосується константних областей, то в легких ланцюгах локалізована один домен, а в важких - три або чотири домена. У підставі розвилки знаходиться шарнірний ділянку, що сприяє обмеженому просторовому переміщенню поліпептидних ланцюгів. При обробці папаїном молекула lg гідролізується в області шарнірного ділянки, утворюючи три фрагмента: два антиген-зв'язуючих FAB (Рис. 30.9) і один, здатний до кристалізації, - Fc. Легкі і важкі ланцюги пов'язані між собою дисульфідними зв'язками.
Мал. 30.9. Кристалічна структура РАВ-фрагмснтов імуноглобуліну G (по PDB-200I) C'hiara, J. В., Strura, Е., Wilson, J .; Science, 264. р. 82, 1994
Теорії сну і сновидіння - вікова анатомія і фізіологія Для пояснення процесу засипання існує кілька теорій. Згідно з одними ( теорія деафферентации ) Сон настає в результаті припинення дії чинників, що підтримують стан неспання. Інші теорії пояснюють наступ сну процесом збудження визначених структур мозку - «Центрів сну», які відповідним чином
Теорії пам'яті - вікова анатомія і фізіологія Існує багато теорій, що пояснюють механізми пам'яті, але безсумнівно, що в їх основі лежать процеси встановлення складних систем зв'язків між нейронами і їх збереження протягом того чи іншого проміжку часу. Следовая теорія пам'яті була сформульована ще І. М. Сеченовим, який пов'язував
Таутомерія і деякі інші фізико-хімічні властивості підстав - біохімія Перераховані пуринові і піримідинові підстави містять пов'язану систему кратних зв'язків і заступники (групи - ОН і -NH 2 ). Зазначені структурні особливості обумовлюють здатність пуринових і піримідинових основ до різних типів таутомерних перетворень: лакто-лактімномудля оксіпроізводних і
Сумісність різних фізіотерапевтичних процедур - сестринський догляд в фізіотерапевтичної практиці Для отримання вираженого клінічного ефекту необхідно слідувати принципам раціонального призначення фізичних методів лікування. На амбулаторному етапі відновного лікування кількість фізіотерапевтичних процедур обмежують. В один день не рекомендується: призначати 2 загальні процедури; послідовно
Ступінь дисоціації (іонізації). Сила електролітів - біохімія людини У розчиніслабких електролітів поряд з іонами існують неіонізовані молекули. Для кількісної характеристики повноти дисоціації введено поняття ступеня дисоціації (іонізації). Ступенем дисоціації (іонізації) електроліту сц, називається відношення числа молекул, що розпалися на іони, до загального
Структури мозку, які беруть участь в організації рухів - вікова фізіологія і психофізіологія Процес організації рухів передбачає існування ієрархічно побудованої системи взаємовідносин між корковими і підкірковими структурами мозку, а також між різними рівнями центральної нервової системи. підкіркові структури : червоне ядро середнього мозку функціонує у взаємодії з кортікоспінальних