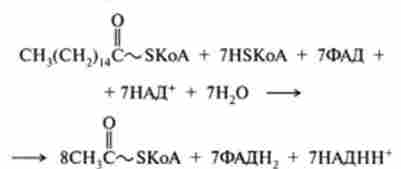
| Головна |
| «« | ЗМІСТ | »» |
|---|
Внутрішня мембрана мітохондрій непроникна для ацил-КоА, що утворився в цитоплазмі. Переносником активованої жирної кислоти є карнітин (у-тріме-
тіламіно-р-гидроксибутират) (CH3)3N-СН2-СН (ОН) СН2-СООН. Це широко поширене з'єднання, особливо багато його в м'язовій тканині. У транспорті ацил-КоА беруть участь фермент - карнітин-ацил- трансфераза (Е,) і транспортний білок (карнітин: ацилкарнітину-транслока- за). Ацил-КоА, з'єднуючись з карнітином, при дії карнітин-ацілтранс- фсрази утворює ацилкарнітину (ефір карнітину і жирної кислоти), який за участю транслокази проникає всередину мітохондрії:
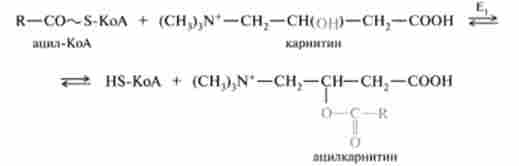
Після проходження ацилкарнітину через мембрану мітохондрії відбувається зворотна реакція - розщеплення ацилкарнітину за участю мітохондріального HS-KoA і карнітин-ацілтрансферази (Е,):
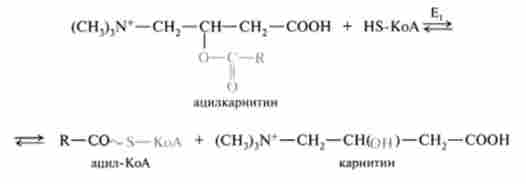
При цьому карнітин повертається в цитоплазму клітини, а ацил-КоА піддається в мітохондріях окислення. Слід зазначити, що карнітин-ацил- трансфсраза є основним регуляторним ферментом процесу окислення жирних кислот. Інгібітором цього ферменту є вихідний интермедиат синтезу жирних кислот - Малоні-КоА. Таким чином, якщо активується липогенез, збільшується концентрація Малоні-КоА, який пригнічує карнітин-ацил-КоА-трансферазу і вимикає р-окислення.
Послідовність реакцій р-окислення ацил-КоА в матриксі мітохондрій. Окислення ацил-КоА, в результаті якого відбувається отщепле-
Про
II
ня двухуглеродний фрагмента СН3З-S-KoA і окислення р-вуглецевого атома кислоти, каталізується чотирма ферментами, відомими під загальною назвою оксидази жирних кислот. Ця система локалізована в матриксі мітохондрій в безпосередній близькості від дихального ланцюга, інтегрованої у внутрішню мембрану мітохондрій. Таким чином, окислення ацил-КоА до ацетил-КоА, в процесі якого відбувається відновлення НАД * і ФАД, пов'язане з синтезом АТФ шляхом окисного фосфорилювання.
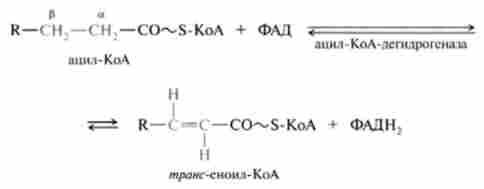
1. Реакція дегидрирования, катализируемая ФАД-залежної ацил-КоА- дегідрогеназ, призводить до утворення а, (3-ненасишенного ацил-SKoA. Фермент має стереоспеціфічность, тому в результаті цієї реакції утворюється тільки транс-ізомер (трш / с-еноіл-КоА):
2. У другій реакції відбувається гідратація ненасиченого транс-tно іл-КоА при дії ферменту еноіл-КоА-гідратази. В результаті утворюється L-P-гідроксіаціл-КоА:

3. Реакція дегидрирования, в процесі якої утворився Р-гідро- ксіаціл-КоА дегидрирующей в P-положення. Цю реакцію каталізує НАД-залежна р-гідроксіаціл-КоА-дегидрогеназа:
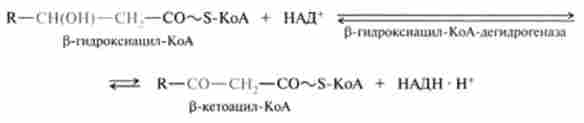
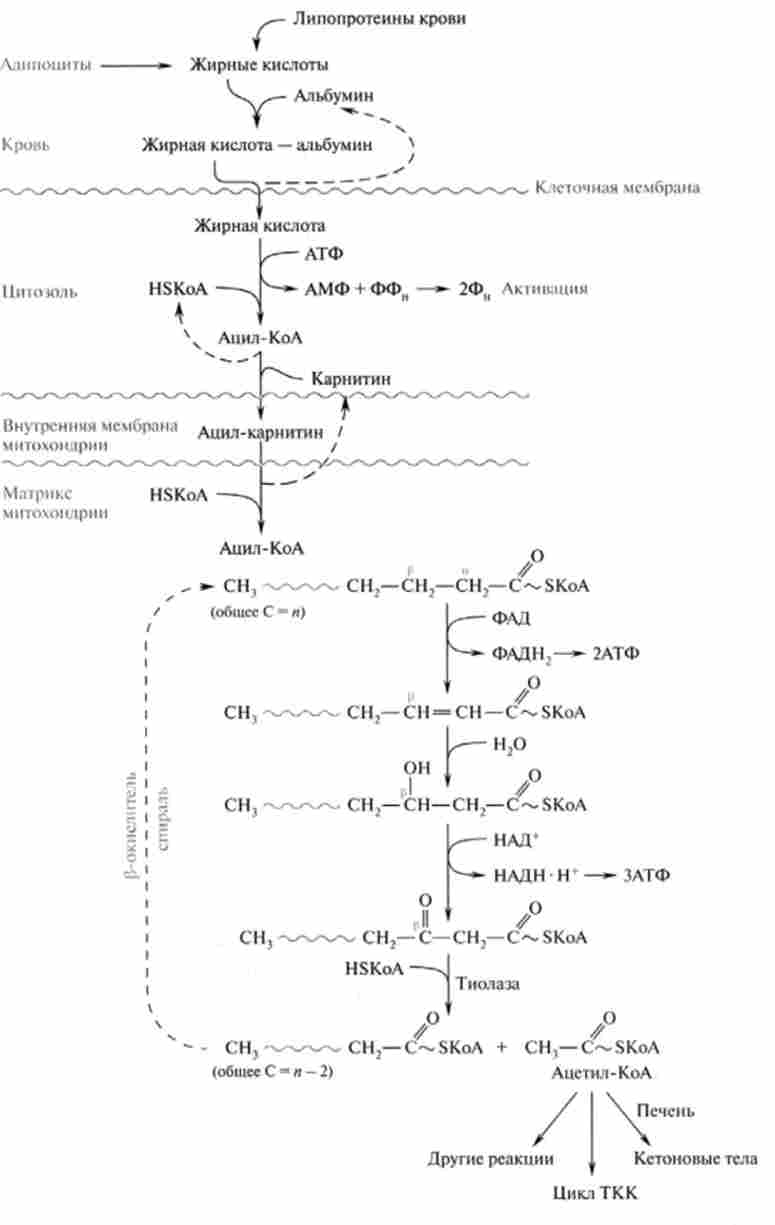
Мал. 23.9. Схема активації і окислення жирних кислот
4. У заключній реакції тіолітічного розщеплення Р-кето ацил-КоА за допомогою ще однієї молекули коензиму А утворюється укорочений на два вуглецевих атома ацил-КоА і двухуглеродний фрагмент у вигляді ацетил-КоА. Реакція каталізується ацетил-КоА-ацілтрансферази (або тіо-
лазой):

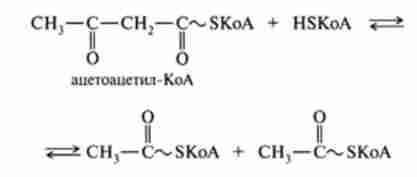
Утворений укорочений ацил-КоА знову вступає в наступний цикл Р-окислення, починаючи з першої реакції дегідрування, і відбувається повторне перетворення цього ацил-КоА в циклі, що складається з чотирьох реакцій, і т. Д. Такий процес р-окислення протікає до освіти четирехуглеродного з'єднання - ацетоацетил-КоА. Остання реакція тіолітіче-
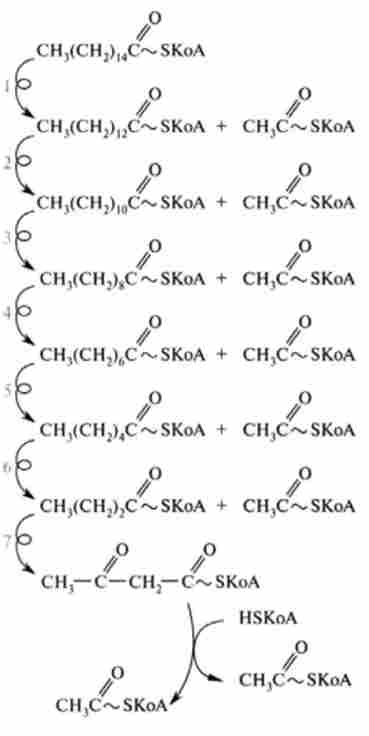
Мал. 23.10. Схема катаболізму пальмітинової кислоти (С16) Шляхом р-окислення:
SLS - позначає повторення чотирьох реакцій Р-окислення;
1-7 - нумерація циклоп Р-окислення
ського розщеплення цього з'єднання призводить до утворення двох молекул ацетил-КоА і тим самим завершує в цілому розпад жирної кислоти за механізмом Р-окислення:
Як зазначалося раніше (гл. 19), молекули ацетил-КоА, що утворилися з жирної кислоти, піддаються повному окисленню до С02 і Н20 в циклі трікар- бонових кислот.
Зі схеми, наведеної на рис. 23.10, видно, що при окисленні однієї молекули
пальмитоил-КоА СН3(СН2)| 4-C ~ SKoA утворилося вісім молекул ацетил-КоА в процесі семи циклів Р-окіс- лення. Отже, сумарне рівняння окислення активованої пальмітинової кислоти можна записати в такий спосіб: