| Головна |
| «« | ЗМІСТ | »» |
|---|
Транспозиції представляють собою переміщення невеликих ділянок генетичного матеріалу в межах однієї хромосоми або між різними хромосомами. Транспозиції відбуваються за участю особливих рухомих або мігруючих генетичних елементів.
Вперше мігруючі генетичні елементи були описані Б. Мак-Клінток в 1947 році у зв'язку з вивченням хромосомних розривів у кукурудзи. Був виявлений мігруючий локус?> $ (диссоциатор), В якому переважно відбуваються розриви хромосом. Сам по собі?> $ Не викликає розривів. Вони з'являються в цьому локусі, якщо в геномі присутня інший мігруючий елемент - Ас {активатор). Обидва ці елементи можуть губитися з частотою кілька відсотків в мейотичному потомство або міняти свою локалізацію при мітотичних поділках. При цьому Ds переміщається тільки в присутності Ас.
впровадження Ds в безпосередній близькості або всередину гена С, контролюючого забарвлення Алейрон насіння, призводило до інактивації гена С, і гетерозиготні насіння С / с / с (нагадаємо, що ендосперм - триплоїдного тканину) виявлялися незабарвленими. У присутності активатора (Ас) діссо- ціатор (Ds) починав переміщатися - іноді залишав локус С. В результаті цього з'являлися пофарбовані плями Алейрон на нефарбованих насінні (рис. 9.8).
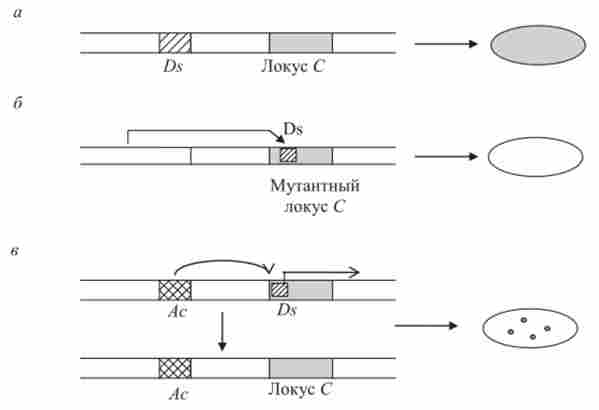
Мал. 9.8. Схема взаємодії активатора (Ас) і диссоциатор ^), інактивована локус С у кукурудзи: а- Оз віддалений від локусу С; б-Ds перемістився в локус З і інактивована його (проявляється як рецесивний аллель с); в-Ас стимулює переміщення /) $, в результаті чого залишає локус С, що виражається в реверсі с-> З в ході мітотичних поділів (поява забарвлених плям на зернах кукурудзи)
Завдяки успіхам генної інженерії в 1980-х рр. вдалося виділити і дослідити Ас, Ds і деякі інші мігруючі елементи кукурудзи (рис. 9.9). Виявилося що Ds - це дефектний делегований варіант Ас. структура елемента Ас виявилася типовою для мігруючих елементів, які до цього часу були вивчені насамперед у бактерій, а також у дрозофіли і дріжджів Басс. сегеч'шае.
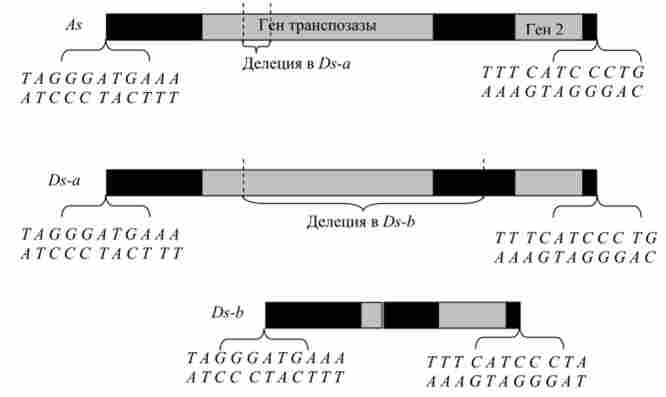
Мал. 9.9. Будова Л * - і Ду-елементів кукурудзи. Показані два гена (світлим), відповідальні за транспозицию, і послідовності інвертованих недосконалих повторів на кінцях
Початок вивченню молекулярної структури мігруючих генетичних елементів поклало відкриття в кінці 1960-х рр. незвичайних мутантів по лактозна оперон Е. coli. У цих мутантів були інактивовані всі три гена / дс-оперона. Частинки бактеріофага А., трансдуцірующего / ос-оперон з таких мутантів, мали надзвичайно високою плавучої щільністю, що вказувало на присутність в lac-оперон мутантів зайвої ДНК. Подібні мутації були знайдені потім і в інших генах Е. coli, фагах X і Р2. Спільними для всіх цих мутантів були инсерции (вставки) більшою чи меншою довжини. Ці вставляються в різні ділянки геному Е. coli молекули ДНК отримали найменування / 5-елементів (від англ, insertion sequences - вставні послідовності).
Розміри / 5-елементів можуть варіювати від 200 до 5 700 п. Н. Все / 5-елементи характеризуються такими структурними особливостями:
4. У точці впровадження кожного / 5-елемента, на його флангах завжди виявляється Дуплікація (в прямій орієнтації) розміром від 4 до 9 п. Н. Ця Дуплікація не є частиною / 5-елемента, а являє собою результат повторень сайту-мішені, в який впроваджується елемент.
зазвичай хромосома Е. coli має кілька / 5-елементів, наприклад 8 копій / 51, 5 копій / 52 і т. д. Вони переміщаються по хромосомі з частотою близько 1 х Ю "6 - 1 х КГ8 на клітинний розподіл. / 5-елементи локалізовані також в / '- факторі Е. coli два / 53, один / 52 і ще один елемент, що позначається уб. Саме за цими мігруючих елементів і відбувається рекомбінація, коли / "- фактор інтегрує з хромосомою Е. coli, утворюючи /// - штами. На це вказують результати вивчення ДНК / '- факторів, у яких ділянка ДНК бактеріальної хромосоми, включений в /' - фактор, виявляється відокремленим від ДНК / '- фактора по обох кінцях однієї і тієї ж послідовністю / 5-елемента. / 5-елементи / '- фактора і такі ж послідовності, розкидані по бактеріальної хромосомі, створюють умови для утворення //// - донорів з різними началами і напрямками перенесення бактеріальної хромосоми.
Міграція / 5-елементів, очевидно, пов'язана з рекомбінацією, проте її механізм відмінний від класичної гомологичной рекомбінації. На це вказує можливість транспозиції навіть в клітинах бактерій, що несуть мутацію гес А, блокує загальну гомологічну рекомбінацію у Е. coli.
Надалі у бактерій були виявлені більш складні мігруючі елементи - транспозони, які відрізняються від / 5-елементів тим, що в них включені деякі гени, що не мають відношення до самого процесу транспозиції. Подібно влаштовані і транспозони еукаріот, наприклад 7> 1 - елемент Sacch. cerevisiae розміром 5 700 п. н., що викликає дуплікації 5 п. н. в точках інтеграції з ДНК хромосом (рис. 9.10). Подібним чином організовані і множинні дисперговані гени (МДГ) D. melanogaster, і ДНК-копії ретровірусів.
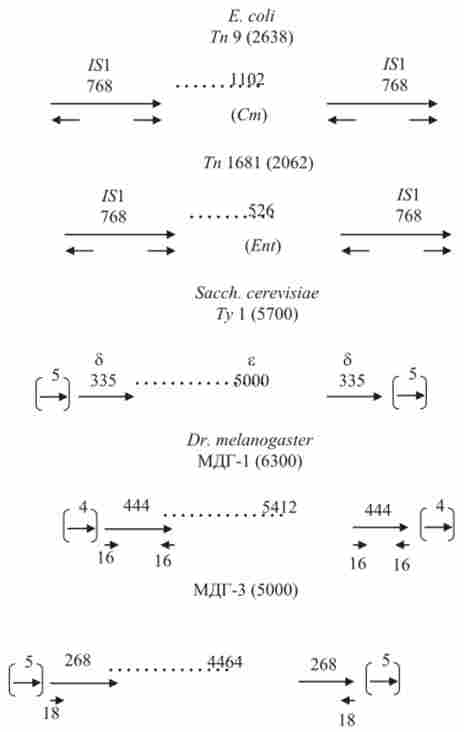
Мал. 9.10. Будова деяких транспозони Е. coli: ТП9 і Тп6' Sacch. cerevisiae: 7> I; D. melanogaster: ПРО-1 і ПРО-3. Цифри - розмір в числі пар нуклеотидів. Стрілки - прямі і інвертовані повтори. Стрілки в дужках - прямий повтор сайту-мішені. Cm - стійкість до хлорамфеніколу; Ent - продукція ентеротоксину; б, е - позначення ділянок ту
Вивчення нуклеотидноїпослідовності дупліціруемих сайтів-мішеней на кінцях мігруючих елементів показало, що вони як правило неоднакові як у розрізняються елементів, так і у одного і того ж елемента, локалізованого в різних місцях. Отже, мігруючі елементи впроваджуються в ті чи інші точки генома незалежно від їх структури. Правда, перевагу в багатьох випадках віддається АТ-містить районам.
Деякі мігруючі елементи, залишаючи точку своєї локалізації, зазнають внутріхромосомную гомологичную рекомбінацію по фланкирующим їх довгим кінцевим повторам. В результаті цього транспозон залишає після себе одну копію свого кінцевого повтору, як це показано, наприклад, для дріжджового транслозона ту (Рис. 9.11), що залишає після себе одну копію 5-повтору. Мабуть, схоже можуть вести себе і МДГ О. melanogaster. Цим пояснюється повторна поява деяких мігруючих елементів в залишених ними сайтах - за рахунок гомологичной рекомбінації з рештою кінцевим елементом.
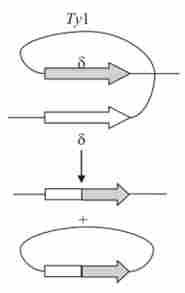
Мал. 9.11. Ексцизія (вирізання) дріжджового транслозона ту за рахунок рекомбінації між термінальними повторами елемента 8
Для транспозони еукаріот розглядають три механізму транспозиції:
Зазвичай переміщення мігруючих елементів не спрямовані і досить рідкісні. Вони відбуваються з частотою 1 х 10-5 - 1 х 10 "8, про що можна судити по частоті ревертірованія мутантів, що виникли в результаті впровадження цих елементів в безпосередній близькості від генів або всередину конкретних генів. У той же час можна створити такі експериментальні ситуації, при яких частота транспозиції різко зростає, а переміщення транспозони перестають бути випадковими. У лініях дрозофіли, тривалий час (більше 500 поколінь) селектировать на зниження статевої активності самців і загальної життєздатності, відбирається невипадковий малюнок розташування МДГ в хромосомах. Про це можна судити по гібридизації клонованих і радіоактивних копій МДГ з ДНК гігантських хромосом D. melanogaster безпосередньо на цитологічних препаратах слинних залоз личинок дрозофіли (In situ). Як тільки селекція в негативну сторону припинялася, відбувалося швидке (протягом декількох поколінь) переміщення копій МДГ, різко змінюється картину їх розташування на хромосомах. Це доводить, що міграція транспозони чуйно відображає напрямок селекції і пов'язана з адаптивною цінністю організму.
На основі цього з'являються бактеріальні Епіс-фактори стійкості, «збирають» по кілька генів, які забезпечують виживання бактерій в період масової терапії антибіотиками.