За аксону відбувається не тільки проведення електричного сигналу, він також є каналом для транспорту різних речовин: білків, медіаторів, низькомолекулярних з'єднань, клітинних органел і т. Д. Речовини всередині аксона можуть рухатися як від тіла клітини до закінчення {Антероградний транспорт), так і від закінчення аксона до тіла клітини (Ретроградний транспорт). Ретроградний транспорт сприяє поверненню речовин в тіло клітини для переробки або розпаду. Він є основним способом переміщення трофічних молекул (фактор росту нерва).
За швидкістю аксоплазматичний транспорт поділяють на швидкий (приблизно 410 мм на добу) і повільний (1-2 мм на добу). Швидкість транспорту залежить від виду яку переносять молекули. Так, структурні білки переміщаються з самої повільною швидкістю (1-2 мм на добу); клітинні органели (мітохондрії, бульбашки і т. п.) рухаються швидше (близько 400 мм на добу). Швидкий транспорт виявлений у всіх нейронах теплокровних тварин, він однаковий в тонких і товстих аксонах, в міелінізірованних і неміе- лінізірованних.
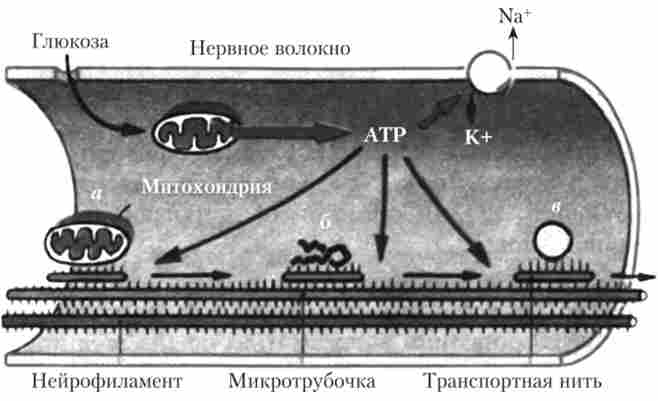
Як пояснення передбачуваного механізму аксонного транспорту пропонується гіпотеза «транспортних ниток» (рис. 4.2). У аксоні є елементи клітинного цитоскелету - білкові нитки: нейрофіламенти, мікротрубочки і так звані транспортні нитки, на яких і розташовується переноситься речовина. Всі ці елементи взаємодіють між собою за типом взаємодії м'язових білків актину і міозину. Під час транспорту відбуваються процеси, аналогічні процесам м'язового скорочення з витратою енергії АТФ (активний процес), і транспортні нитки як би ковзають уздовж елементів клітинного цитоскелету, переносячи таким чином різні молекули і органели.
Антероградний транспорт здійснюється білком кінезин, а ретроградний - білком динеина. Специфічні рецептори на поверхні органел обумовлюють приєднання до них або кінезин, або діне- ина і таким чином визначають напрямок руху органел.
Мал. 4.2. Гіпотетичний механізм аксонного транспорту1
Автономна (вегетативна) нервова система - анатомія центральної нервової системи В результаті вивчення даного розділу студент повинен: знати відмінні риси ВНС в порівнянні з іншими нейроанатомічсскімі і функціональними блоками мозку; принципи поділу ВНС на симпатичну, парасимпатичну і метасім- патичної складові; їх взаємодія; центральні і периферичні компоненти ВНС; їх
Ацілгліцероли - біохімія ацілгліцероли, або нейтральні ліпіди, - найбільш поширена в природі група ліпідів. Ці сполуки являють собою складні ефіри жирних кислот і трехатомного спирту гліперола (гліцериди), в якому можуть бути етерифіковані одна, дві або три гідроксильні групи гліцс- рола з утворенням соответствен
Аспекти обміну ліпідів в відновної медицини - біохімія людини В організм з їжею повинні поступати незамінні жирні кислоти. При тривалому їх відсутності в їжі у тварин спостерігається відставання в рості, розвиваються характерні ураження шкіри і волосяного покриву. Описані випадки недостатності незамінних жирних кислот і у людини. Гак, у дітей грудного
Анестетики - стоматологія. Ендодонтія Ін'єкційна анестезія вимагає введення анестетика в тканини. Амідні анестетики представлені 0,25% -ним, 0,5% -ним, 1,0% -ним, 2,0% -ним розчинами лідокаїну або ксілокаіна, 2,0% -ним розчином тримекаина, 3,0% -ним розчином мепівакаіна або скандонеста, 4% -ним розчином артикаина (ультракаїн,
Амінокислоти і пептиди в промисловості та медицині - біохімія Щорічно в світі виробляється більше 200 тис. Тонн амінокислот, які використовуються в основному як харчові добавки і компоненти кормів для худоби. Традиційним промисловим методом їх отримання є ферментація, однак все більшого значення набувають хімічні і особливо ферментативні методи синтезу
Активація жирної кислоти в цитоплазмі клітини - біохімія частина 2. Реакції окислення жирних кислот відбуваються тільки після перетворення її в активовану високоенергетичну форму - ацил-КоА. Цей процес вимагає витрати однієї молекули АТФ, присутності коензиму А і іонів Mg2 +; каталізує перетворення вільної жирної кислоти в який активується форму - фермент