| Головна |
| «« | ЗМІСТ | »» |
|---|
Глікоген - основна форма депонування вуглеводів у тварин - синтезується головним чином в печінці, складаючи до 6% від маси печінки, і в м'язах, де його зміст рідко перевищує 1%.
Глікоген печінки виконує важливу функцію в підтримці фізіологічної концентрації глюкози в крові, перш за все в проміжках між прийомами їжі. Функція м'язовогоглікогену полягає в тому, що він є доступним джерелом глюкози в самому м'язі. Глікоген локалізована в цитоплазмі клітин у формі гранул, які крім глікогену містять ферменти, що беруть участь в його обміні.
Слід звернути увагу, що розпад і синтез глікогену катализируются різними ферментами і, отже, протікають по різним метаболічним шляхам.
Синтез глікогену починається через 1-2 години після прийому їжі, що містить вуглеводи. Процес синтезу глікогену вимагає витрати енергії АТФ.

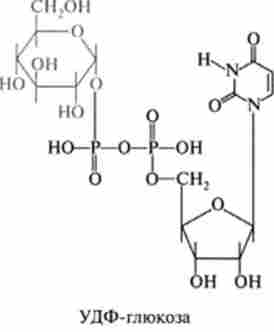
3. Новоутворена фосфорілірованний глюкоза вже безпосередньо втягується в синтез глікогену. Однак попередньо вона взаємодіє з УТФ, і при дії ферменту глюкозо-1 - фосфатурідінтрансферази (інша назва УДФГ-пірофосфорілаза) утворюється урідіндіфосфатглюкози (УДФ-глюкоза):

Структурна формула УДФ-глюкози:
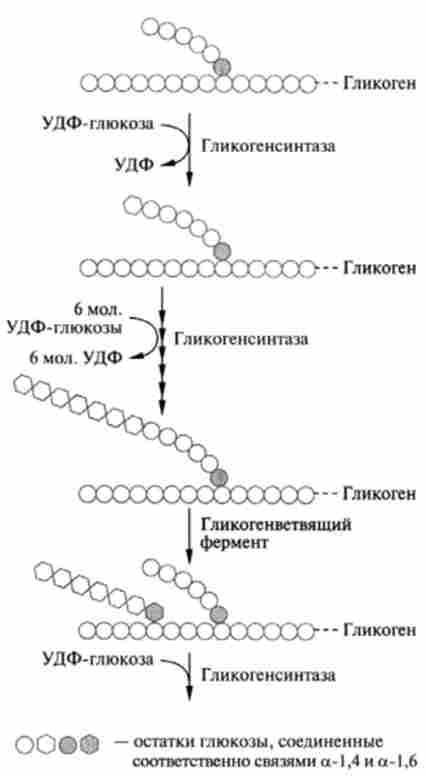
Мал. 20.5. синтез глікогену
Новоутворена УДФ-глкжоза є переносником і донором активованих глюкозільних залишків в подальшій ферментативної реакції синтезу глікогену. Ця функція нуклеозіддіфосфатсахаров була встановлена аргентинським біохіміком Л. Лелуара, удостоєним Нобелівської премії за ці роботи.
4. Реакція, що приводить до утворення глікогену, відбувається при перенесенні глюкозного залишку, що входить до складу УДФ-глюкози, на глико- зідную «затравочную» ланцюг глікогену.
При цьому утворюється а (1-> 4) -гліко- зідная зв'язок між першим атомом вуглецю, який додається залишку глюкози і 4-гідроксильною групою залишку глюкози в ланцюзі глікогену. Ця реакція каталізується ферментом глікоген- синтазою (рис. 20.5).

Таким чином, в результаті цієї реакції відбувається тільки подовження ланцюга, т. Е. Вона вимагає присутності поліглюкозной «затравки»: самого глікогену, амілози, амілопектину або будь-якого олігосахариду з довжиною ланцюга не менше чотирьох глюкозних залишків і призводить до утворення лінійного полімеру а -1- * 4-глюкана.
У рослин донором глюкозільних груп при синтезі крохмалю служить АДФ-Е-глюкоза, а не УДФ-проізводнис (гл. 16).
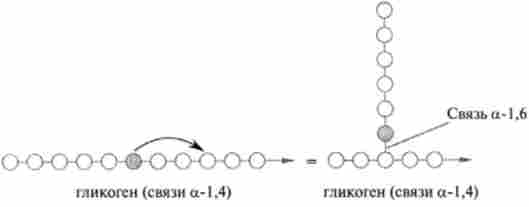
Розгалуження ланцюгів глікогену в результаті утворення а-1? 6-зв'язків (по одній на кожні 8-12 залишків, з'єднаних а-1? 4-зв'язками) каталізується іншим ферментом - а-глюкан-ветвящегося глюкозілтрансферазой (відомої також під назвою «гл ікогенветві щий фермент»). Цей фермент відщеплює невеликі фрагменти ланцюга 1,4-глюкана (шість або сім мономерних одиниць) і переносить їх на ту ж саму (або іншу аналогічну) ланцюг, але в положення 6, в результаті чого утворюється 1,6-зв'язок за схемою:
Регуляція глікогеногенеза. У гл. 18 приведена регуляція розщеплення глікогену (глікогенолізу) за допомогою оборотної ковалентного хімічного модифікації ферменту глікогснфосфорілази (фосфорилювання - дефосфорілірованіе). Глікогенсінтази також існує в двох формах - фосфо рілірованной і дефосфорілірованном, але вона регулюється реціпропно по відношенню до глікогенфосфорилази, т. Е. Прямо протилежним чином. В результаті складного каскаду реакцій фосфорилювання активної гли- когенсінтази а призводить до переходу її в фосфорильовану неактивну форму:
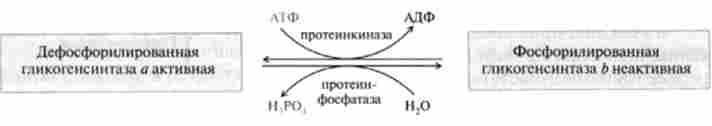
Протсінкіназа і протєїнфосфатаза - це тс ж самі ферменти, які брали участь у взаимопревращении а- і 6-форм глікогенфосфорі- лази.
Таким чином, такі гормони, як адреналін і глюкагон, дія яких опосередковано цАМФ, синхронно пригнічують синтез глікогену і активують глікогеноліз, тим самим їх гормональний вплив призведе до підвищення цукру в клітинах печінки і крові (рис. 20.6).
Слід помститися, що в м'язовій тканині рецептори глюкагону відсутні і регуляторний дію цього гормону на обмін глікогену відзначено лише в печінці.
Відома також аллостерічеськая регуляція активності глікогснсінтази Ь. Будучи фосфорілірованним, цей фермент мало або повністю неактивний, однак глюкозо-6-фосфат (при високій концентрації) по аллостерічному механізму значною мірою підвищує активність глікогенсінтази. Ця форма глікогенсінтази називається D-формою або залежною (dependent) формою від присутності глюкозо-6-фосфату. а дефосфорілірованном форма - активної і під час відсутності глюкозо-6-фосфату - 1-формою або незалежної (independent) від присутності цього модулятора.
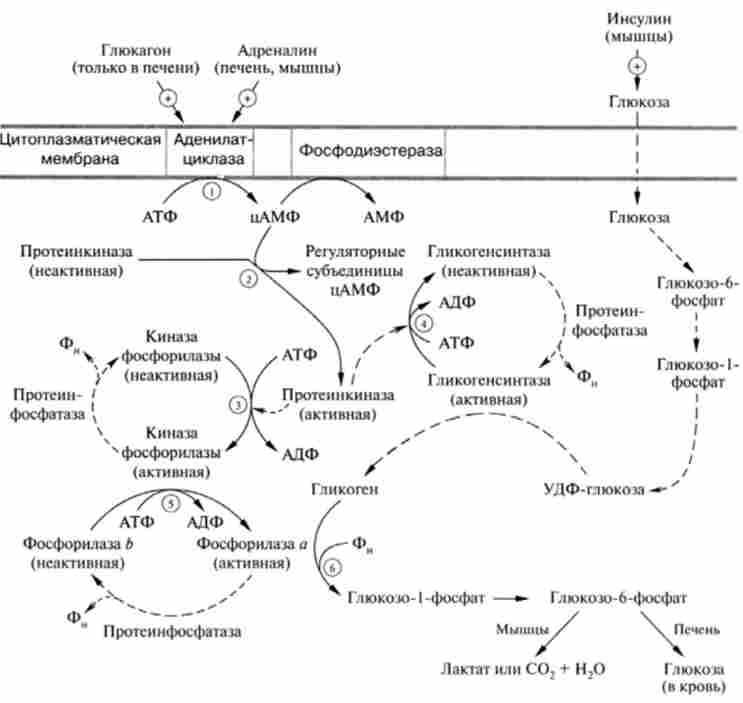
Мал. 20.6. Гормональна регуляція синтезу і деградації глікогену:
(Т) - (б) - каскад реакцій післядії глюкагону і адреналіну (суцільна лінія); стимулюючу дію інсуліну на синтез глікогену (пунктирна лінія)
Підвищення продуктивності лікарських на синтез глікогену в м'язах надає також інсулін, сприяючи дефосфорілірованіе глікогенсінтази за рахунок активації протєїнфосфатаза, що каталізує реакцію дефосфорилирования цього ферменту.