| Головна |
| «« | ЗМІСТ | »» |
|---|
Клітини багатоклітинних організмів, як тварин, так і рослинних, відокремлені від свого оточення оболонкою. Клітинна оболонка, або плазмалемма, тваринних клітин утворена мембраною, покритий зовні шаром гликокаликса товщиною 10-20 нм. Основними складовими гликокаликса служать комплекси полісахаридів з білками (глікопротеїни) і жирами (гліколіпіди). Зсередини до мембрани примикає кортикальний (корковий) шар цитоплазми товщиною 0,1-0,5 мкм, в якому невідомі рибосоми і бульбашки, але в значній кількості знаходяться мікротрубочки і мікрофіламенти, що мають в своєму складі скоротні білки.
Плазмалемма виконує відмежовує, бар'єрну, транспортну і рецепторну функції. Завдяки властивості виборчої проникності вона регулює хімічний склад внутрішнього середовища клітини. У плазмалемме розміщені молекули рецепторів, які вибірково розпізнають певні біологічно активні речовини (гормони). В утриманні (заякоріванню) цих речовин на клітинної поверхні беруть участь білки кортикального шару. Наявність в оболонці рецепторів дає клітинам можливість сприймати сигнали ззовні, щоб доцільно реагувати на зміни в навколишньому середовищі або стані організму. У пластах і прошарках сусідні клітини утримуються завдяки наявності різного виду контактів, які представлені ділянками плазма- леми, що мають особливу будову.
У клітці виділяють ядро і цитоплазму. клітинне ядро складається з оболонки, ядерного соку, ядерця і хроматину. функціональна роль ядерної оболонки полягає у відокремленні генетичного матеріалу (хромосом) еукаріотичної клітини від цитоплазми з притаманними їй численними метаболічними реакціями, а також регуляції двосторонніх взаємодій ядра і цитоплазми. Ядерна оболонка складається з двох мембран, розділених околоя- дерну (перінуклеарним) простором. Останнє може сполучатися з канальцями цитоплазматичної мережі.
Ядерна оболонка пронизана порами діаметром 80-90 нм. Область пори або порові комплекс з діаметром близько 120 нм має певну будову, що вказує на складний механізм регуляції ядерно-цитоплазматичних переміщень речовин і структур. Кількість пір залежить від функціонального стану клітини. Чим вище синтетична активність в клітці, тим більше їх число. Підраховано, що у нижчих хребетних тварин в ері- тробластах, де інтенсивно утворюється і накопичується гемоглобін, на 1 мкм2 ядерної оболонки доводиться близько 30 пір. У зрілих еритроцитах названих тварин, які зберігали ядра, на 1 мкм2 оболонки залишається до п'яти пір, т. е. в 6 разів менше.
В області порового комплексу розташовується так звана щільна пластинка - білковий шар, що підстилає на всьому протязі внутрішню мембрану ядерної оболонки. Ця структура виконує насамперед опорну функцію, так як при її наявності форма ядра зберігається навіть в разі руйнування обох мембран ядерної оболонки. Припускають також, що закономірний зв'язок з речовиною щільної пластинки сприяє впорядкованого розташування хромосом в інтерфазних ядрі.
основу ядерного соку, або матриксу, складають білки. Ядерний сік утворює внутрішнє середовище ядра, в зв'язку з чим він грає важливу роль в забезпеченні нормального функціонування генетичного матеріалу. У складі ядерного соку присутні нитчасті, або фіблярні, білки, з якими пов'язане виконання опорної функції: в матриксі знаходяться також первинні продукти транскрипції генетичної інформації - гетероядерні РНК (гяРНК), які тут же піддаються процесингу, перетворюючись в мРНК (див. розд. 3.4.3.2).
Ядро являє собою структуру, в якій відбувається утворення і дозрівання рібосомал'них РНК (рРНК). Гени рРНК займають певні ділянки (в залежності від виду тварини) однієї або декількох хромосом (у людини 13-15 і 21-22 пари) - ядерцеві організатори, в області яких і утворюються ядерця. Такі ділянки в метафазних хромосомах виглядають як звуження і називаються вторинними перетяжками. За допомогою електронного мікроскопа в полісом виявляють нитчастий і зернистий компоненти. Нитчастий (фібрилярний) компонент представлений комплексами білка і гігантських молекул РНК-попередниць, з яких потім утворюються більш дрібні молекули зрілих рРНК. В процесі дозрівання фібрили перетворюються в рібонуклеопротеіновие зерна (гранули), якими представлений зернистий компонент.
Хроматіновие структури у вигляді грудочок, розсіяних в нуклео- плазмі, є інтерфазних формою існування хромосом клітини (див. розд. 3.5.2.2).
В цитоплазмі розрізняють основну речовину (матрикс, гіалоплазма), включення і органели. Основна речовина цитоплазми заповнює простір між плазмалеммой, ядерної оболонкою і іншими внутрішньоклітинними структурами. Звичайний електронний мікроскоп не виявляються в ньому будь-якої внутрішньої організації. Білковий склад гіалоплазми різноманітний. Найважливіші з білків представлені ферментами гліколізу, обміну цукрів, азотистих основ, амінокислот і ліпідів. Ряд білків гіалоплазми служить субодиницями, з яких складаються таких структур, як мікротрубочки.
Основна речовина цитоплазми утворює справжню внутрішню середу клітини, яка об'єднує всі внутрішньоклітинні структури і забезпечує взаємодію їх один з одним. Виконання матриксом об'єднує, а також каркасної функції може бути пов'язано з виявляється за допомогою надпотужного електронного мікроскопа мікротрабекулярной мережі, утвореної тонкими фибриллами товщиною 2-3 нм і пронизує всю цитоплазму. Через гіалоплазму здійснюється значний обсяг внутрішньоклітинних переміщень речовин і структур. Основна речовина цитоплазми слід розглядати так само, як складну колоїдну систему, здатну переходити з золеобразного (рідкого) стану в гелевидний. В процесі таких переходів відбувається робота.
Про функціональне значення таких переходів см. Розд. 2.3.8.
включеннями (Рис. 2.5) називають відносно непостійні компоненти цитоплазми, які служать запасними поживними речовинами (жир, глікоген), продуктами, такими, що підлягають виведенню з клітини (гранули секрету), а також деякі пігменти.
органели - це постійні структури цитоплазми, що виконують в клітці життєво важливі функції.
виділяють органели загального значення і спеціальні. Останні в значній кількості присутні в клітинах, спеціалізованих до виконання певної функції, але в незначній кількості можуть зустрічатися і в інших типах клітин. До них відносять, наприклад, микроворсинки всмоктуючої поверхні епітеліальної клітини кишечника, вії епітелію трахеї і бронхів, синаптичні пухирці, що транспортують речовини - переносники нервового збудження з однієї нервової клітини на іншу або клітку робочого органу, міофібрили, від яких залежить скорочення м'язи. Детальний розгляд спеціальних органел входить в завдання курсу гістології.
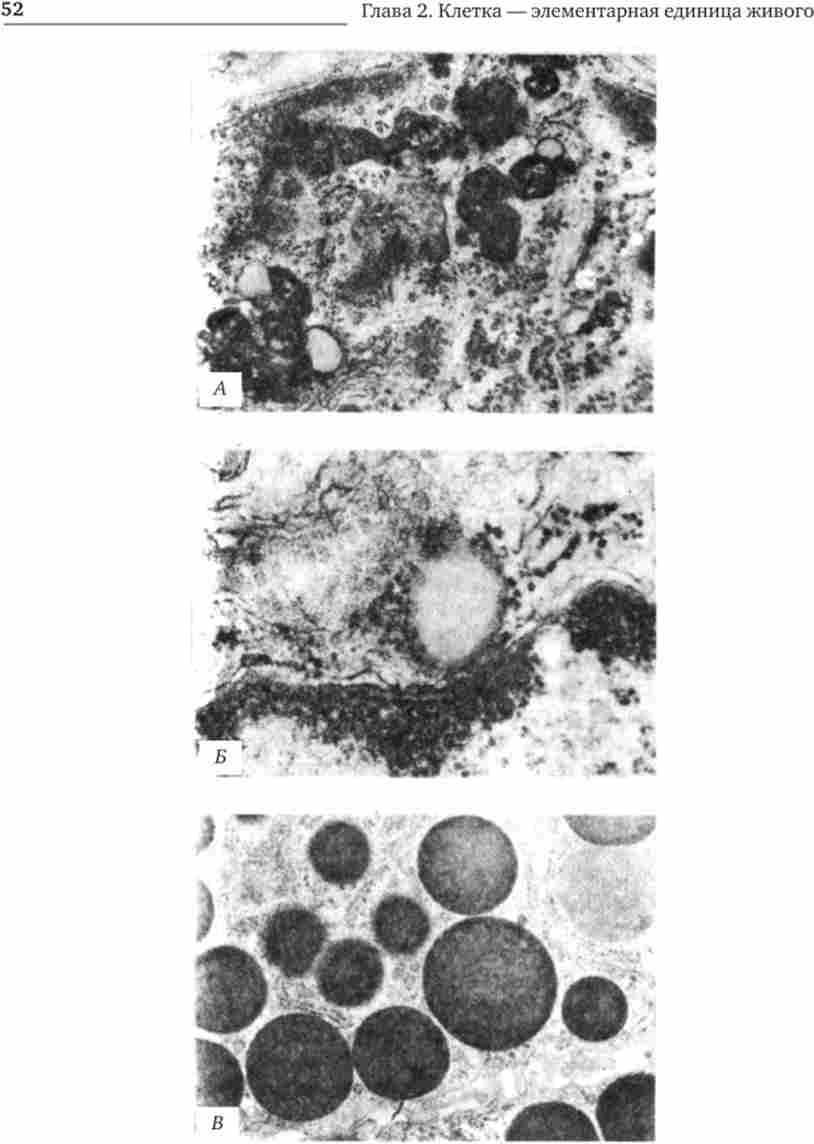
Мал. 2.5. Цитоплазматичні включення еукаріотичної клітини.
А - липофусцин; Б - ліпідна крапля; В - гранули секрету
До органел загального значення відносять елементи канальцевої і вакуолярної системи у вигляді шорсткою і гладкою цитоплазматичної мережі, пластинчастий комплекс, мітохондрії, рибосоми і полісоми, лізосоми, пероксисоми, мікрофібрили і мікротрубочки, центріолі клітинного центру. У рослинних клітинах виділяють також хлоропласти, в яких відбувається фотосинтез.
канальцева і вакуолярна системи утворені сполученими або окремими трубчастими або сплощеними (цистерна) порожнинами, обмеженими мембранами і поширюються по всій цитоплазмі клітини. Нерідко цистерни мають бульбашковидні розширення. У названій системі виділяють шорстку і гладку цитоплазматичну мережі (Див. Рис. 2.3). Особливість будови шорсткою мережі полягає в прикріпленні до її мембран полісом.
В силу цього вона виконує функцію синтезу певної категорії білків, переважно видаляються з клітки, наприклад секретується клітинами залоз. В області шорсткою мережі відбувається утворення білків і ліпідів цитоплазматичних мембран, а також їх складання. Щільно упаковані в шарувату структуру цистерни шорсткою мережі є ділянками найбільш активного білкового синтезу і називаються ергастоплазма.
Мембрани гладкої цитоплазматичної мережі позбавлені полісом. Функціонально ця мережа пов'язана з обміном вуглеводів, жирів і інших речовин небілкової природи, наприклад стероїдних гормонів (в статевих залозах, кірковому шарі надниркових залоз). За канальцям і цистерн відбувається переміщення речовин, зокрема секре- тіруемого залозистої кліткою матеріалу, від місця синтезу в зону упаковки в гранули. У ділянках печінкових клітин, багатих структурами гладкою мережі, руйнуються і знешкоджуються шкідливі токсичні речовини, деякі ліки (барбітурати). У бульбашках і канальцях гладкою мережі поперечно-смугастої мускулатури зберігаються (депонуються) іони кальцію, які відіграють важливу роль в процесі скорочення.
рибосома - це округла рібонуклеопротеіновая частка діаметром 20-30 нм. Вона складається з малої і великої субодиниць, об'єднання яких відбувається в присутності матричної (інформаційної) РНК (мРНК). Одна молекула мРНК зазвичай об'єднує кілька рибосом на зразок нитки бус. Таку структуру називають полисомой. Полісоми вільно розташовуються в основній речовині цитоплазми або прикріплені до мембран шорсткою цитоплазматичної мережі. В обох випадках вони служать місцем активного синтезу білка. Порівняння співвідношення кількості вільних і прикріплених до мембран полісом в ембріональних недиференційованих і пухлинних клітинах, з одного боку, і в спеціалізованих клітинах дорослого організму - з іншого, призвело до висновку, що на полісомах гіалоплазми утворюються білки для власних потреб (для «домашнього» користування) даної клітини, тоді як на полісомах гранулярной мережі синтезуються білки, що виводяться з клітки і використовуються на потреби організму (наприклад, травні ферменти, білки грудного молока).
Пластинчастий комплекс Гольджі утворений сукупністю дик- тіосом числом від кількох десятків (зазвичай близько 20) до кількох сотень і навіть тисяч на клітину.
діктіосома (Рис. 2.6, Л) представлена стопкою з 3-12 сплощених дископодібних цистерн, від країв яких отшнуровива- ються бульбашки (везикули). Обмежені певною ділянкою (локальні) розширення цистерн дають більші бульбашки (вакуолі). У диференційованих клітинах хребетних тварин і людини діктіосоми зазвичай зібрані в околоядерной зоні цитоплазми. У пластинчастому комплексі утворюються секреторні пухирці або вакуолі, вміст яких складають білки та інші сполуки, що підлягають виведенню з клітини. При цьому попередник секрету (просекрет), що надходить в диктиосом із зони синтезу, піддається в ній деяких хімічних перетворень. Він також відокремлюється (сегрегуючий) у вигляді «порцій», які тут же одягаються мембранної оболонкою. У пластинчастому комплексі утворюються лізосоми. У діктіосоми синтезуються полісахариди, а також їх комплекси з білками (глікопротеїни) і жирами (гліколіпіди), які входять до складу глікокаліксу клітинної оболонки.
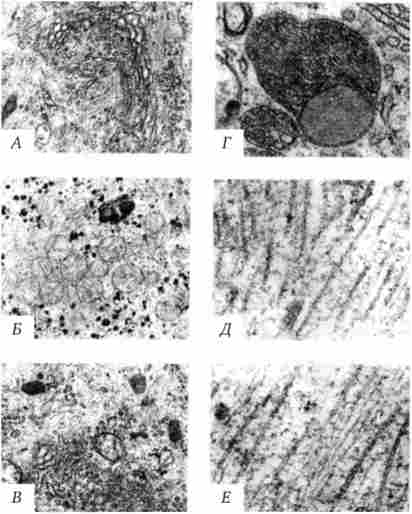
Мал. 2.6. Загальноклітинні органели. Пояснення см. В тексті
мітохондрії (Рис. 2.6, Б) - це структури округлої або палочковидной, нерідко ветвящейся форми товщиною 0,5 мкм і довжиною зазвичай до 5-10 мкм. У більшості тварин клітин кількість мітохондрій коливається від 150 до 1500, проте в жіночих статевих клітинах їх число досягає декількох сотень тисяч. У сперматозоїдах нерідко присутній одна гігантська мітохондрія, спірально закручена навколо осьової частини джгутика. Одна розгалужена мітохондрія виявлена в клітці такого паразита людини, як трипаносома.
Оболонка мітохондрії складається з двох мембран, що розрізняються за хімічним складом, набору ферментів і функцій. Внутрішня мембрана утворює впячивания листоподібною (Крісті) або трубчастої (Тубуль) форми. Простір, обмежений внутрішньою мембраною, становить матрикс органели. У ньому за допомогою електронного мікроскопа виявляються зерна діаметром 20-40 нм. Вони накопичують іони кальцію і магнію, а також полісахариди, наприклад глікоген.
У матриксі розміщений власний апарат біосинтезу білка органели. Він представлений 2-6 копіями кільцевої і позбавленою гістонів (як у прокаріотів) молекули ДНК, рибосомами, набором транспортних РНК (тРНК), ферментами редуплікаціі ДНК, транскрипції і трансляції спадкової інформації. За основними властивостями: розмірами та структурою рибосом, організації власного спадкового матеріалу - цей апарат схожий з таким у прокаріотів і відрізняється від апарату біосинтезу білка цитоплазми еукаріотичної клітини (чим підтверджується симбиотична гіпотеза походження мітохондрії; см. Розд. 1.5). Гени власної ДНК кодують нуклеотидні послідовності мітохондріальних рРНК і тРНК, а також послідовності амінокислот деяких білків органели, головним чином її внутрішньої мембрани. Амінокислотні послідовності (первинна структура) більшості білків мітохондрії закодовані в ДНК клітинного ядра і утворюються поза органели в цитоплазмі.
Головна функція мітохондрії складається в ферментативном витягу з певних хімічних речовин енергії (шляхом їх окислення) і накопиченні енергії в біологічно використовуваної формі (шляхом синтезу молекул аденозинтрифосфату - АТФ). В цілому цей процес називається окислювальним фосфорилюванням. В енергетичній функції мітохондрії беруть активну участь компоненти матриксу і внутрішня мембрана. Саме з цією мембраною пов'язані ланцюг перенесення електронів (окислення) і АТФ-синтетаза, що каталізує поєднане з окисленням фосфорилювання АДФ в АТФ. Серед побічних функцій мітохондрії можна назвати участь в синтезі стероїдних гормонів і деяких амінокислот (глутамінова).
лізосоми (Рис. 2.6, В) являють собою бульбашки діаметром зазвичай 0,2-0,4 мкм, які містять набір ферментів кислих гідролаз, які каталізують при низьких значеннях pH гидролитичне (у водному середовищі) розщеплення нуклеїнових кислот, білків, жирів, полісахаридів. Їх оболонка утворена одинарної мембраною, покритої іноді зовні волокнистих білковим шаром (на електронограммах «облямовані» бульбашки). Функція лізо- сом - внутрішньоклітинний перетравлювання різних хімічних сполук і структур.
первинними лизосомами (Діаметр 100 нм) називають неактивні органели, вторинними - органели, в яких відбувається процес перетравлення. Вторинні лізосоми утворюються з первинних. Вони підрозділяються на гетеролізосоми (Фаголізосоми) і аутолі- зосоми (Цитолізосоми). У перших (рис. 2.6, Г) перетравлюється матеріал, що надходить в клітку ззовні шляхом піноцитозу і фагоцитозу, по-друге руйнуються власні структури клітини, які завершили свою функцію.
Мікротільця складають збірну групу органел. Це обмежені однією мембраною бульбашки діаметром 0,1-1,5 мкм з дрібнозернистим матриксом і нерідко кристалоїдними або аморфними білковими включеннями. До цієї групи відносять, зокрема, перок- сісоми. Вони містять ферменти оксидази, що каталізують утворення пероксиду водню, який, будучи токсичним, руйнується потім під дією ферменту пероксидази. Ці реакції включені в різні метаболічні цикли, наприклад в обмін сечової кислоти в клітинах печінки і нирок. В печінковій клітині число пероксисом досягає 70-100.
До органел загального значення відносять також деякі постійні структури цитоплазми, позбавлені мембран. микротрубочки (Рис. 2.6, Д) - трубчасті утворення різної довжини із зовнішнім діаметром 24 нм, шириною просвіту 15 нм і товщиною стінки близько 5 нм. Зустрічаються у вільному стані в цитоплазмі клітин або як структурні елементи джгутиків, війок, митотичного веретена, центриолей. Вільні мікротрубочки і мікротрубочки війок, джгутиків і центриолей мають різну стійкість до руйнуючих дій, наприклад хімічним (колхіцин). Микротрубочки будуються з стереотипних субодиниць білкової природи шляхом їх полімеризації. У живій клітині процеси полімеризації протікають одночасно з процесами деполімеризації. Співвідношенням цих процесів визначається кількість мікротрубочок. У вільному стані мікротрубочки виконують опорну функцію, визначаючи форму клітин, а також є факторами спрямованого переміщення внутрішньоклітинних компонентів.
микрофиламентами (Рис. 2.6, Е) Називають довгі тонкі освіти, іноді утворюють пучки і виявляються по всій цитоплазмі. Існує кілька різних типів микрофиламентов. актинові мікрофіламенти завдяки присутності в них скоротних білків (актину) розглядають в якості структур, що забезпечують клітинні форми руху, наприклад амебоидние. Їм приписують також каркасну роль і участь в організації внутрішньоклітинних переміщень органел і ділянок гіалоплазми.
По периферії клітин під плазмалеммой, а також в околоядерной зоні виявляються пучки мікрофіламентів товщиною 10 нм - проміжні філаменти. В епітеліальних, нервових, гліальних, м'язових клітинах, фибробластах вони побудовані з різних білків. Проміжні філаменти виконують, по-видимому, механічну, каркасну функцію.
Актинові мікрофібрили і проміжні філаменти, як і мікротрубочки, побудовані з субодиниць. В силу цього їх кількість залежить від співвідношення процесів полімеризації і деполімеризації.
Для тварин клітин, частини клітин рослин, грибів і водоростей характерний клітинний центр, до складу якого входять цен- тріолі. Центріол' має вигляд «порожнього» циліндра діаметром близько 150 нм і довжиною 300-500 нм. Її стінка утворена 27 мікротрубочками, згрупованими в 9 триплетів. У функцію центріолей входить освіту ниток мітотичного веретена, які також утворені мікротрубочками. Центриоли поляризують процес поділу клітини, забезпечуючи розбіжність сестринських хроматид (хромосом) в анафазе мітозу.