| Головна |
| «« | ЗМІСТ | »» |
|---|
Процеси регуляції вуглеводного обміну знаходяться під контролем великої кількості різних факторів, які забезпечують координацію і збалансованість складних хімічних процесів розпаду і синтезу вуглеводів.
Існує два основних рівня контролю:
Однак незалежно від рівня регуляції регуляторний сигнал реалізується через внутрішньоклітинні механізми, т. Е. Шляхом зміни активності або кількості ферменту або швидкості трансмембранного перенесення речовин (метаболічна регуляція).
На активність ферментів вуглеводного обміну впливають концентрації субстратів і продуктів реакції, кисневий режим, виборча проникність біомембран, концентрація коферментів та ін. (Гл. 18 і 19).
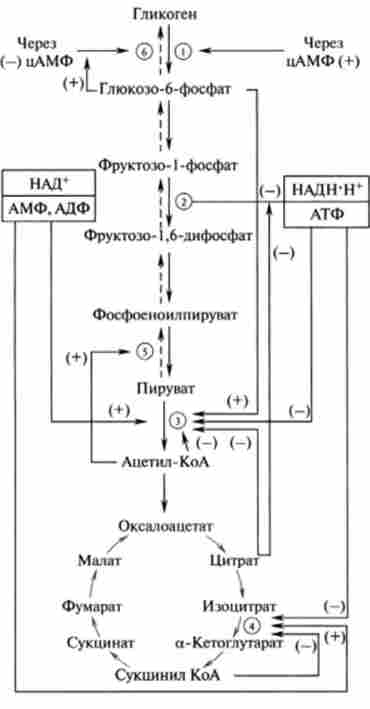
Мал. 20.7. Схема регуляторних механізмів ряду процесів вуглеводного обміну ссавців:
реакції катаболізму (глікогеноліз, гліколіз, окисне декарбоксилювання пірувату. цикл ТКК) - суцільні лінії; реакції анаболізму (глюконеогенез, синтез глікогену) - пунктирні лінії. Активація ферментів (+); інгібування (-). Головні регуляторні ферменти: (Т) - глікогенфосфорилази; @ - Фос фофруктокнназа: (з) - піруватдекарбоксилази; 0- ізоіітратдегідрогеназа; 0- піруваткарбо- ксілаза; 0- глікогенсінтази
В основі регуляторних механізмів зміни активності ферментів вуглеводного обміну лежать два шляхи: аллостерічеськая регуляція за типом прямого і зворотного зв'язку і оборотна ковалентная модифікація ферментів
(Фосфорилювання - дефосфорілірованіе). Раніше вже розглядалися регуляторні механізми різних процесів метаболізму вуглеводів. На рис. 20.7 наведена узагальнена схема механізмів регуляції вуглеводного обміну в організмі ссавців, в тому числі і гормонального (через цАМФ). У схемі приведені тільки основні пункти контролю.
Як видно з рис. 20.7, можна виділити три групи регуляторних сигналів:
Підвищення рівнів АДФ, АМФ. НАД+ представляє сигнал, що збільшує швидкість катаболізму і зменшує швидкість анаболізму, в той час як високі рівні АТФ, НАДН (первинне джерело енергії для синтезу АТФ) представляють собою сигнали, що знижують швидкість реакцій катаболізму. Всі впливу за типом прямого і зворотного зв'язку узгоджені один з одним і з впливом АТФ, АДФ, АМФ, НАД *, НАДН. Слід пам'ятати, що в організмі людини, вищих тварин на всіх стадіях синтезу і розпаду вуглеводів регуляція вуглеводного обміну здійснюється за участю ЦНС і гормонів. Механізми цього рівня регуляції докладно розглянуті в гл. 13.
Регуляція метаболізму вуглеводів здійснюється гормонами підшлункової залози - інсуліном і Глюка гон ом; гормонами коркового шару надниркових залоз - глюкокортикоїдами.
Інсулін є єдиним гормоном, різко знижує вміст цукру в крові. Його дія на вуглеводний обмін поліфункціональної. Основні механізми регуляції пов'язані з підвищенням в присутності інсуліну проникності клітинних мембран для транспорту глюкози всередину клітини, а також опосередковано через активацію синтезу регуляторних ферментів катаболізму глюкози - гексокінази і фосфофруктокинази, ферменту синтезу глікогену - глікогенсінтази (гл. 13).
Адреналін і глюкагон здійснюють регуляцію метаболізму глікогену шляхом зміни активності глікогенфосфорилази і глікогенсінтази (через цАМФ) таким чином, що гальмування глікогеногенеза і стимуляція глико- геноліза здійснюються одночасно, т. Е. Реціпропно. Глюкокортикоїди (11-гідроксістероід) підсилюють глюконеогенез за рахунок інтенсифікації катаболізму білків і амінокислот в тканинах і залучення проміжних метаболітів в процес глюконеогенезу. Таким чином, в розглянутих випадках адреналін, глюкагон, глюкокортикоїди діють як антагоністи інсуліну. На вміст цукру в крові впливає також гормон щитовидної залози тироксин (подібно до інсуліну). Гормони передньої долі гіпофіза - гормон росту (соматотропін), АКТГ і, ймовірно, інші чинники підвищують рівень цукру в крові, однак механізми дії цих гормонів в значній мірі є опосередкованими, оскільки вони стимулюють мобілізацію з жирової тканини вільних жирних кислот, які є інгібіторами споживання глюкози.