| Головна |
| «« | ЗМІСТ | »» |
|---|
Принципово важливою властивістю генетичної інформації є її здатність до переносу (передачі) як в межах однієї клітини, так і від батьківської клітини до дочірнім або між клітинами різних індивідуумів в процесах клітинного ділення і розмноження організмів. Що стосується напрямків внутрішньоклітинного перенесення генетичної інформації, то в разі ДНК-містять організмів вони пов'язані з процесами реплікації молекул ДНК, т. Е. З копіюванням інформації, або з синтезом молекул РНК (транскрипцією) і утворенням поліпептидів
(Трансляцією) (рис. 5.15). Кожен із зазначених процесів здійснюється на основі принципів матричної і комплементарності.
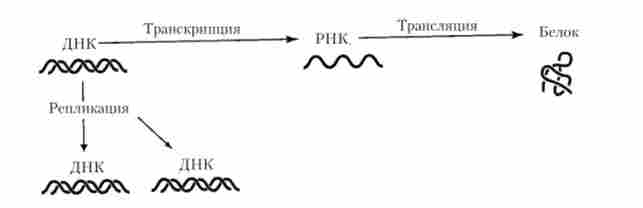
Мал. 5.15. Основні напрямки внутрішньоклітинного перенесення генетичної інформації
Сформовані уявлення про перенесення генетичної інформації за схемою ДНК -? РНК - + білок прийнято називати «центральної догмою» молекулярної біології. Поряд з цим (найбільш поширеним) напрямком перенесення, який іноді позначають як «загальний перенос», відома й інша форма реалізації генетичної інформації ( «спеціалізований перенесення»), виявлена у РНК-вірусів. У цьому випадку спостерігається процес, який отримав назву зворотної транскрипціїу при якому первинний генетичний матеріал (вірусна РНК), що проник в клітку-господаря, служить матрицею для синтезу комплементарної ДНК за допомогою ферменту зворотної транскриптази {Ревертази)у кодируемой вірусним геномом. Надалі можлива реалізація інформації синтезованої вірусної ДНК в звичайному напрямку. Отже, спеціалізований перенесення генетичної інформації здійснюється за схемою РНК> ДНК> РНК? білок.
Транскрипція - перший етап загального перенесення генетичної інформації, являє собою процес біосинтезу молекул РНК за програмою ДНК. Принциповий сенс цього процесу полягає в тому, що інформація структурного гена (або декількох розташованих поруч генів), записана в формі нуклеотидноїпослідовності кодує нитки ДНК в орієнтації 3'5 переписується (транскрибується) в нуклеотидну послідовність молекули РНК, що синтезується в напрямку 5 * -? З * на основі комплементарного відповідності дезоксирибонуклеотидов матричної нитки ДНК риб про нуклеотидам РНК (A-У, Г-Ц, Т-А, Ц-Г) (рис. 5.16). Як продуктів транскрипції (транскриптов) можна розглядати всі типи молекул РНК, що беруть участь в біосинтезі білків в клітині: матричні (інформаційні) РНК (мРНК, або іРНК), рібосом- ні РНК (рРНК), транспортні РНК (тРНК), малі ядерні РНК (мяРНК).
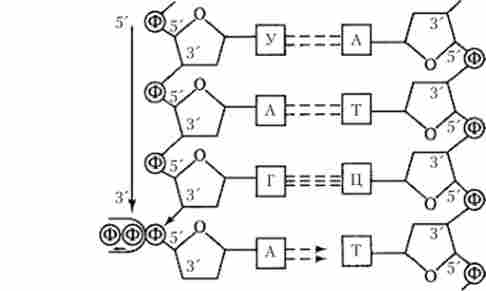
Мал. 5.16. Синтез молекули РНК на матричної нитки ДНК (стрілкою показано напрямок, в якому йде зростання ланцюга РНК)
Процес транскрипції забезпечується комплексною дією ряду ферментів, до числа яких належить РНК-полімераза, що представляє собою складний білок, що складається з декількох субодиниць і здатний виконувати кілька функцій. На відміну від прокаріотів (бактерій), в клітинах яких є РНК-полімераза лише одного типу, що забезпечує синтез різних молекул РНК, у еукаріот встановлено наявність ядерних РНК-полімерази трьох типів (I, II, III), а також РНК-полімерази клітинних органел , що містять ДНК (мітохондрій, пластид). РНК-полімераза I знаходиться в полісом і бере участь в синтезі більшості молекул рРНК, РНК-полімераза II забезпечує синтез мРНК і мяРНК, а РНК-полімераза III здійснює синтез тРНК і одного варіанта молекул рРНК - 5SPHK.
Транскрипція підрозділяється на три основні стадії: ініціацію (початок синтезу РНК), елонгацію (подовження полинуклеотидной ланцюжка) і терминацию (закінчення процесу).
ініціація транскрипції залежить від попереднього специфічного зв'язування РНК-полімерази з впізнаваною нею короткою нуклеотидной послідовністю в ділянці молекули ДНК (промоторі), розташованому перед стартовою точкою структурного гена, з якої починається синтез РНК. Промотори різних структурних генів можуть бути ідентичними або містять відрізняються один від одного послідовності нуклеотидів, що, ймовірно, визначає ефективність транскрибування окремих генів і можливості регуляції самого процесу транскрипції. Промотори багатьох генів прокаріотів мають в своєму складі універсальну послідовність 5-ТАТААТ-З '(блок Прібнова), яка розташовується перед стартовою точкою на відстані близько 10 нуклеотидів і розпізнається
РНК-полімераза. Інша щодо часто зустрічається впізнавана послідовність прокаріотів (5-ТТГ. АЦА-3 ') зазвичай виявляється на відстані приблизно 35 нуклеотидів від стартової точки. У геномах еукаріот функцію впізнавання для РНК-полімерази II можуть виконувати універсальні послідовності ТАТА (блок Хогнесса), ЦААТ і складаються з повторюваних нуклеотидів Г і Ц (ГЦ-мотиви). При цьому та чи інша промоторних область може містити або одну із зазначених послідовностей або комбінацію двох або трьох таких послідовностей.
Специфічне міцне зв'язування РНК-полімерази з тим чи іншим впізнаваним нею ділянкою промоторной області дозволяє їй почати процес розплітання молекули ДНК аж до стартової точки, з якої вона починає здійснювати полімеризацію рибонуклеотидов з використанням в якості матриці однонітевую 3'-5'-фрагмента ДНК.
Елонгація. Подальше розплітання ДНК структурного гена супроводжується подовженням синтезованого полирибонуклеотидов (елонгацією нитки РНК), що триває аж до досягнення РНК-полімераза області термінатора.
Терминация - процес припинення реплікації ДНК, яка відбувається за допомогою термінатора. Останній являє собою нуклеотидну послідовність ДНК, яка впізнається РНК-полімераза за участю інших білкових факторів термінації, що призводить до закінчення синтезу транскрипта і його від'єднання від матриці. У більшості випадків термінатор знаходиться в кінці структурного гена, забезпечуючи синтез однієї моногенной молекули мРНК. При цьому у прокаріотів можливий синтез полігенною молекули мРНК, що кодує синтез двох і більше полі- пептидних ланцюжків. Відбувається безперервне транскрибування декількох розташованих поруч один з одним структурних генів, що мають один загальний термінатор. Полигенная мРНК може містити в своєму складі нетрансльовані міжгенних області (спейссри), що розділяють кодують ділянки для окремих поліпептидів, що, ймовірно, забезпечує подальше розділення і самих синтезованих поліпептидів.
Оскільки структурні гени еукаріот мають переривчасте (мозаїчне) будова, їх транскрипція має специфічні особливості, що відрізняють її від транскрипції у прокаріот. У разі еукаріотичного гена, що кодує синтез поліпептиду, цей процес починається з транскрибування всієї нуклеотидної послідовності, що містить як екзон- ні, так і інтронів ділянки ДНК. Новоутворена при цьому молекула мРНК, що відображає структуру всього музичного гена, яку називають гетерогенної ядерної РНК (гяРНК) або про-матричної РНК (про-мРНК), зазнає потім процес дозрівання (процесинг мРНК).
Процесинг полягає в ферментативному розрізуванні первинного транскрипту (гяРНК) з подальшим видаленням його інтронних ділянок і возз'єднанням (сплайсингом) екзонних ділянок, які формують безперервну послідовність, що кодує зрілої мРНК, яка в подальшому бере участь в трансляції генетичної інформації. Як приклад можна розглянути схему процесингу мРНК, що синтезується при транскрипції гена р-глобіновой ланцюжка (рис. 5.17).
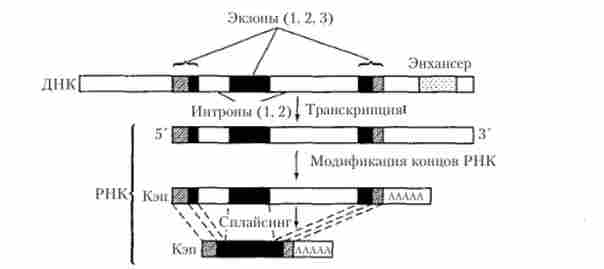
Мал. 5.17. Процесинг мРНК р-глобинового гена людини
У процессинге беруть участь і короткі молекули мяРНК, що складаються приблизно з 100 нуклеотидів, які представляють собою послідовності, які є комплементарними послідовностями на кінцях інтронних ділянок гяРНК. Парування комплементарних нуклеотидів мяРНК і гяРНК сприяє згортанню в петлю інтронних ділянок і зближенню відповідних екзонних ділянок гяРНК, що, в свою чергу, робить їх доступними розрізати дії ферментів (нуклеаз). Отже, молекули мяРНК забезпечують правильність вирізання інтронів з гяРНК.
Під час процесингу відбувається також модифікація 5 - і З'-решт формується зрілої молекули мРНК. Принциповий сенс цього процесу можна розглянути на схемах процесингу гена р-глобіну людини (рис. 5.17) і повної нуклеотидної послідовності зрілої мРНК, що утворюється в результаті цього процесу (рис. 5.18).

Мал. 5.18. Нуклеотидная послідовність зрілої мРНК р-глобинового гена людини
На 5'-кінці послідовності (рис. 5.18) є короткий не- трансльований (лідируючий) ділянку, що складається з 17 кодонів, які марковані цифрами зі знаком «мінус». Ця ділянка кодується транскрибируемой (але нетрансльовані) областю першого екзона р-гена (заштрихована на рис. 5.17). Модифікація цієї ділянки полягає в освіті 5'-кінцевого кепа (від англ, cap - ковпачок, шапочка), що представляє собою залишок 7-мстився | -уанозіна, приєднаний до сусіднього нуклеотиду незвичайним способом (за допомогою трифосфатних зв'язку). Передбачається, що основна функція кепа пов'язана з впізнавання специфічної послідовності молекули рРНК, що входить до складу рибосоми, що забезпечує точне прикріплення всього лідируючого ділянки молекули мРНК до певної ділянки цієї рибосоми і ініціацію процесу трансляції. Можливо також, що кеп оберігає зрілу мРНК від передчасного ферментативного руйнування під час її транспортування з ядра в цитоплазму клітини.
Нуклеотидная послідовність зрілої мРНК р-глобинового гена людини починається з 7-метілгуанозіна на 5'-кінці (кеп-сайт), за яким слідує короткий нетрансльовані ділянку РНК. Перший транслюється кодон (АУТ) виділено шрифтом і позначений цифрою 0, оскільки кодується їм амінокислота (метіонін) надалі вищепляются з поліпептиду (першої амінокислотою зрілого білка буде валін, який кодується ГУГ). Виділено також стоп-кодон УАА (кодон 147), на якому закінчується трансляція (поліпептид складається з 146 амінокислот), і сигнальна послідовність для поліаденілювання (ААУААА) на -кінці.
Модифікація -кінця мРНК Р-глобіну, також має коротку нетрансльовані послідовність, кодованих відповідної областю третього екзона p-гена (рис. 5.17), пов'язана з утворенням полі- аденілового (поли А) «Хвоста» молекули, що складається з 100-200 послідовно з'єднаних залишків адениловой кислоти. Для дії ферменту, що здійснює поліаденілювання, нс потрібна матриця, але потрібна присутність на -кінці мРНК сигнальної послідовності ААУААА (рис. 5.18). Передбачається, що поліаденіловой «хвіст» забезпечує транспорт зрілої мРНК до рибосоми, захищаючи її від ферментативного руйнування, але сам поступово руйнується ферментами цитоплазми, відщеплюється один за іншим кінцеві нуклеотиди.
трансляція як черговий етап реалізації генетичної інформації полягає в синтезі поліпептиду на рибосомі, при якому в якості матриці використовується молекула мРНК (зчитування інформації в напрямку 5е-? З *) - Слід зауважити, що в клітинах прокаріотів, які не мають справжнього ядра з оболонкою, хромосомний генетичний матеріал (ДНК) знаходиться в цитоплазмі, що визначає безперервний характер взаємозв'язку процесів транскрипції і трансляції. Іншими словами, що утворився лідируючий 5-кінець молекули мРНК, синтез якої ще не завершено, вже здатний вступати в контакт з рибосомою, ініціюючи синтез поліпептиду, т. Е. Транскрипція і трансляція йдуть одночасно. Що стосується еукаріот, то процеси транскрипції їх ядерної генетичної інформації і її трансляції повинні бути розділені в часі в зв'язку з процессингом молекул РНК і необхідністю їх подальшої упаковки і транспортування з каріоплазми в цитоплазму за участю спеціальних транспортних білків.
Як і в разі транскрипції, процес трансляції можна умовно поділити на три основні стадії: ініціацію, елонгацію і терминацию.
для ініціації трансляції принципово важливе значення має специфічність структурної організації групи ідентичних рибосом (полірібосоми, або полісоми), яка може брати участь у синтезі первинної структури певної білкової молекули (поліпептиду), кодируемой відповідної мРНК. Окрема рибосома є клітинну органеллу, що складається з молекул рРНК, які визначають її специфічність, і з білків. У складі рибосоми є дві структурні субодиниці (велика і мала), Які можна диференціювати на підставі їх здатності по-різному осідати при ультрацентріфугірованіі препаратів очищених рибосом із зруйнованих клітин, т. Е. За коефіцієнтом седиментації (величиною 5). При певних умовах в клітці може відбуватися розділення (дисоціація) цих двох субодиниць або їх об'єднання (асоціація).
Рибосоми прокаріотів, а також мітохондрій і хлоропластів складаються з великої та малої субодиниць з величинами 505 і 305 відповідно, тоді як у еукаріот ці субодиниці мають інші розміри (605 і 405). Оскільки процес трансляції більш детально було досліджено у бактерій, то найчастіше його розглядають у зв'язку зі структурою рибосом цих організмів. Рибосома містить дві ділянки (рис. 5.19), що мають пряме відношення до ініціації трансляції, позначених як А-ділянку (аміноацільний) і P-ділянка (пептідільний), специфічність яких визначається поєднанням відповідних областей субодиниць 505 і 305. При дисоціації субодиниць рибосоми ці ділянки стають «недобудованими», що призводить до зміни їх функціональної специфічності.
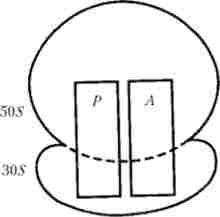
Мал. 5.19. Будова бактеріальної рибосоми:
Р - пептідільний ділянку; А - аміноацільний ділянку
У процесі трансляції беруть участь також молекули тРНК, функції яких полягають у транспортуванні амінокислот з цитозолю (цитоплазматичного розчину) до рибосом. Молекула тРНК, що має вторинну структуру у формі «конюшини», містить трійку нуклеотидів (антикодон), яка забезпечує її комплементарное з'єднання з відповідним кодоном (кодоном) молекули мРНК, що кодує синтез поліпептиду на рибосомі, і акцепторні ділянку (на -кінці молекули ), до якого приєднується певна амінокислота (див. рис. 5.6). Процес приєднання кожної з 20 амінокислот до акцепторному кінця відповідної тРНК пов'язаний з її активацією певним варіантом ферменту аміноацил-тРНК-синтетази з використанням енергії аденозінтріфос- фатов (молекул АТФ). Утворився при цьому специфічний комплекс тРНК і амінокислоти, який отримав назву аміноацил-тРНК, переміщається потім до рибосоми і бере участь в синтезі поліпептиду.
Ініціація трансляції забезпечується точним з'єднанням лідируючого 5-кінця молекули мРНК з певною областю малої субодиниці диссоциированной рибосоми таким чином, що в «недобудованому» P-ділянці виявляється стартовий (який ініціює) кодон АУТ цієї молекули (рис. 5.20, а). Функціональна особливість такого Р-ділянки полягає в тому, що він може бути зайнятий тільки ініціює аміноацил тРНК з антикодоном УАЦ, яка у еукаріот несе амінокислоту метіонін, у бактерій - формілметіонін. Оскільки синтез поліпептиду завжди починається з Анконца і нарастаег в напрямку до С-кінця, то все білкові молекули, синтезовані в клітинах прокаріотів, повинні починатися з N-формілметіонін, а у еукаріот - з TV-метіоніну. Однак в подальшому ці амінокислоти ферментативно вищепляются під час процесингу білкової молекули (див. Рис. 5.18).
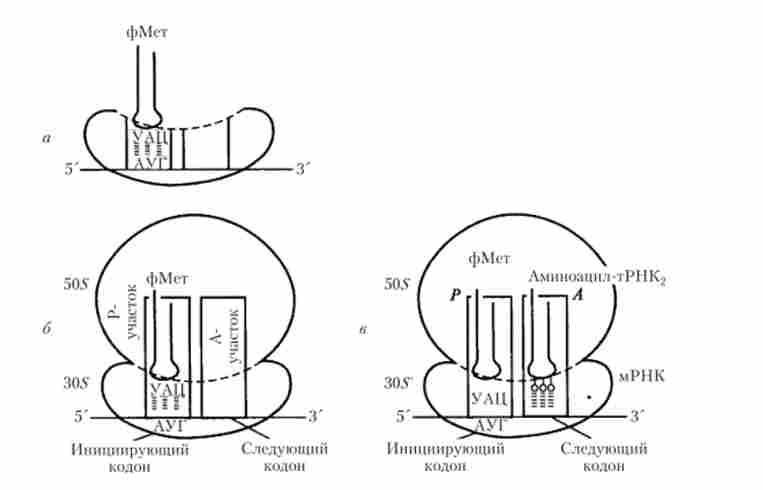
Мал. 5.20. Початкові етапи трансляції: а - ініціює комплекс; б, в- елонгація
Після утворення ініціюючого комплексу в «недобудованому» Л-ділянці (рис. 5.20) стає можливим возз'єднання малої і великої субодиниць рибосоми, що призводить до «добудування» Р-ділянки і Л-ділянки. Лише після цього наступна аміноацил-тРНК може займати / 4-ділянку на основі принципу комплементарності її антикодону відповідного кодону мРНК, що знаходиться в цій ділянці (рис. 5.20, б, в).
процес елонгації починається з утворення пептидного зв'язку між ініціює (першої в ланцюжку) і подальшої (другий) амінокислотами. Потім відбувається переміщення рибосоми на один триплет мРНК в напрямку 5'? З ', супроводжуване від'єднанням ініціює тРНК від матриці (мРНК), від ініціації амінокислоти і виходом її в цитоплазму. При цьому друга за рахунком аміноацил-тРНК пересувається з Л-ділянки в Р-ділянку, а звільнився / 4-ділянку займається наступної (третьої за рахунком) аміноацил-тРНК. Процес послідовного пересування рибосоми «триплетними кроками» по нитці мРНК повторюється, супроводжуючись звільненням тРНК, що надходять в P-ділянка, і нарощуванням амінокислотноїпослідовності синтезованого поліпептиду.
Терминация трансляції пов'язана з входженням одного з трьох відомих стоп-кодонів мРНК в / 4-ділянку рибосоми. Оскільки такий триплет не несе інформації про будь-яку амінокислоті, але пізнається відповідними білками термінації, то процес синтезу поліпептиду припиняється і він від'єднується від матриці (мРНК).
Після виходу з функціонуючої рибосоми вільний 5-кінець мРНК може вступати в контакт з наступною рибосомою полісомной групи, ініціюючи синтез ще одного (ідентичного) поліпептиду. Отже, розглянутий рибосомні цикл послідовно повторюється за участю кількох рибосом однієї і тієї ж полісоми, в результаті чого синтезується група ідентичних поліпептидів.
Посттранашціонная модифікація поліпептиду являє собою завершальний етап реалізації генетичної інформації в клітині, що призводить до перетворення синтезованого поліпептиду в функціонально активну молекулу білка. При цьому первинний поліпептид може зазнавати процесинг, що складається в ферментативном видаленні ініціюють амінокислот, відщепленні інших (непотрібних) амінокислотних залишків і в хімічній модифікації окремих амінокислот. Потім відбуваються процес згортання лінійної структури поліпептиду за рахунок утворення додаткових зв'язків між окремими амінокислотами і формування вторинної структури білкової молекули (рис. 5.21). На цій основі формується ще більш складна третинна структура молекули.
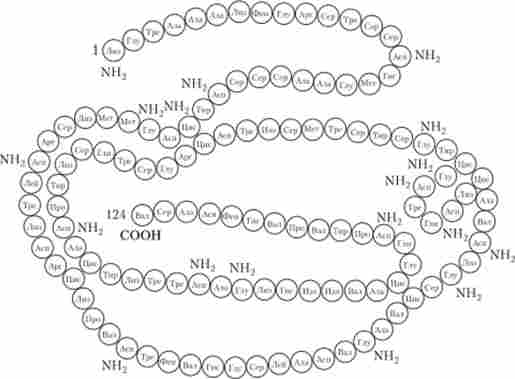
Мал. 5.21. Вторинна структура молекули ферменту рібонуклсази
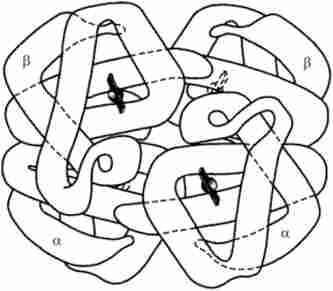
Мал. 5.22. Четвертичная структура молекули гемоглобіну людини
У разі білкових молекул, що складаються більш ніж з одного поліпептиду, відбувається утворення комплексної четвертичной структури, в якій об'єднуються третинні структури окремих поліпептидів. Як приклад можна розглянути модель молекули гемоглобіну людини (рис. 5.22), що складається з двох а-цепочск і двох (J-ланцюжків, які формують стабільну тетрамерную структуру за допомогою водневих зв'язків.
Кожна з глобінових ланцюжків містить також молекулу гема, який в комплексі з залізом здатний зв'язувати молекули кисню, забезпечуючи їх транспортування еритроцитами крові.