| Головна |
| «« | ЗМІСТ | »» |
|---|
До органів розмноження самців тварин відносять насінники з придатками, семяпроводи, сечостатевої канал, додаткові статеві залози і статевий член. Статеві органи самців иннервируются гілками поперекового і крижового сплетінь, а також симпатичної ланцюжка. В іннервації сім'яників бере участь також блукаючий нерв.
Насінники - парні статеві залози, в яких утворюються статеві клітини - спермії (сперматозоїди) і синтезується чоловічий статевий гормон - тестостерон та ін. Зазвичай насінники тварин яйцевидної форми, їх довжина коливається від 11 до 15 см в залежності від виду тварини. Маса сім'яників бика близько 300 ... 350 г (кожен), барана - 200 ... 300, козла - 145 ... 150, жеребця - близько 200, кнура - 250 ... 350 м Насінники покриті серозною і соединительнотканой ( білкову) оболонкою. Радіальні перегородки розділяють тіла насінники на окремі камери, паренхіма яких складається з звивистих сім'яних канальців і сполучної тканини. Під соединительнотканой оболонкою канальців розташовуються фолікулярні клітини (клітини Сертолі) і клітини сперматогенного епітелію, що дають початок чоловічим статевим клітинам.
Під час ембріонального розвитку насінники закладаються в поперекової частини черевної порожнини плода. В останні місяці ембріогенезу або безпосередньо після народження вони опускаються в пахову область і потім в мошонку самця.
Мошонка є мішок зі шкіри, внутрішню поверхню якого вистилають м'язової-еластична і загальна піхвова оболонки. Шкіра мошонки містить велику кількість добре розвинених сальних і потових залоз. Температура в мошонці нижче, ніж в черевній порожнині, що сприяє процесам освіти чоловічих статевих клітин. У разі затримки насінників в черевній порожнині (крипторхізмі) самці повністю втрачають здатність до запліднення (при двосторонньому крипторхізмі) або ж вона дещо знижується (при односторонньому крипторхізмі). Це підтверджує фізіологічну роль мошонки в механізмах терморегуляції сім'яників.
Існує певний механізм підтримки постійного рівня температури в мошонці незалежно від умов зовнішнього середовища. У спекотну погоду м'язової-еластична оболонка і м'язів-подниматель насінники розслабляються, мошонка збільшується в розмірах, і насінники опускаються вниз, т. Е. Теплооот- дача збільшується. Цьому сприяє секреція потових залоз. У холодну погоду мускул-подниматель насінники і мускульноеластічна оболонка скорочуються, насінники підтягуються вгору і наближаються до тіла тварини. Шкіра мошонки зморщується, стінки її товщають, а її поверхня зменшується, що значно знижує тепловіддачу. Посилена секреція сальних залоз також обумовлює стабільність температури. Так, при перепадах температури навколишнього повітря від 15 до 38 ° С температура всередині сім'яників у бика змінюється тільки на 2,3 ° С. Вимкнення функції потових і сальних залоз при забрудненні мошонки або шкірних ураженнях призводить до порушення сперматогенезу.
На процес терморегуляції в сім'яниках також впливає їх кровопостачання. Внутрішня насіннєва артерія починається від аорти в поперековій області і по всій довжині до паховому каналу сильно звивається, що поряд з порівняно невеликим діаметром артерії сприяє охолодженню артеріальної крові. В області пахового каналу артерія його тісний зв'язок з масивним венозних сплетінням внутрішньої насіннєвої вени, в результаті чого створюються сприятливі умови для охолодження артеріальної крові. У спекотну погоду приплив крові до сім'яників послаблюється, а в холодну посилюється.
Гематотестикулярний бар'єр - спеціальний функціональний механізм, що захищає статеві клітини від зовнішніх впливів; за ступенем проникності можна порівняти з гематоенцефалічний. Завдяки цьому бар'єру існують певні відмінності в складі плазми крові і насіннєвий плазми. У насіннєвий плазмі концентрація калію в 2-3 рази вище, ніж в плазмі крові, а кальцію і магнію приблизно в два рази нижче; відсутні глюкоза і фруктоза, але відзначено високий вміст инозитола; рівень білка, в тому числі і імуноглобулінів, значно нижче (близько 5%), ніж в плазмі крові. Основу гематотестикулярного бар'єру складають структури, розташовані між просвітом капілярів і просвітом насіннєвихканальців: ендотелій капілярів, лімфатичні судини інтерстицію, власна оболонка сім'яних канальців, клітини Сертолі, інтерстиціальна тканина сім'яника і фіброзна оболонка сім'яника.
Важлива роль в процесах нормального функціонування сім'яників в цілому і безпосередньо сперматогенного епітелію, який і дає початок чоловічим статевим клітинам, належить фолікулярним клітинам Сертоли. Клітини Сертолі входять до складу епітелію сім'яних канальців, їх циліндричне підстава спочиває на базальній мембрані, а відростки, звернені в бік просвіту сім'яних канальців, утворюють своєрідні контакти з клітинами сперматогенного епітелію. Цитоплазма клітин Сертолі тягнеться від власної оболонки сім'яного канальця до його просвіту і містить значну кількість гладкого і шорсткого ретикулума; в цитоплазмі численні лізосоми здійснюють перетравлення дегенерують статевих клітин. Крім участі в освіті гематотестикулярного бар'єру клітини Сертолі беруть участь в харчуванні і підтримуванні клітин сперматогенного епітелію. У клітинах Сертолі відбувається утворення так званого андрогенсвязивающего білка, що взаємодіє з андрогенами, які необхідні для регуляції процесів сперматогенезу, т. Е. Освіти гаплоїдних чоловічих статевих клітин з диплоїдні первинних статевих клітин. Андрогенсвязивающій білок виробляється під впливом фоллікулстімулірующего гормону (ФСГ) гіпофіза, для якого клітини Сертолі служать мішенями. Клітини Сертолі, в свою чергу, беруть участь в регуляції вироблення фоллікулстімулірующего гормону: вони виробляють речовину, що гальмує секрецію ФСГ і зване інгибіном ФСГ.
Сперматогенез і спермиогенез. Процес утворення чоловічих статевих клітин - сперміїв, або сперматозоїдів, відбувається в стінках сім'яних канальців. У міру розвитку статеві клітини переміщаються від оболонки сім'яного канальця до його центру.
Базальна мембрана звивистих сім'яних канальців вистелена клітинами, що мають диплоїдний набір хромосом - спермато- гоніямі. В результаті мітотичного поділу сперматогонії дають початок первинним сперматоцітов. Первинні сперматоці- ти зазнають мейотичне розподіл, в результаті якого утворюються два вторинних сперматоцита, що дають початок чотирьом гаплоїдний сперматіди. Сперматиди після ряду структурнофункционального перетворень перетворюються в зрілі сперматозоїди або сперміну. Цей процес носить назву сперміогенеза.
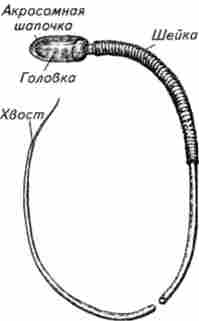
Мал. 9.1. Схема будови спермия ссавців
Спермін - це закінчили своє зростання і розвиток чоловічі гамети, функція яких полягає в заплідненні жіночої статевої клітини при заплідненні. У ссавців спермій складається з головки, шийки, тіла і хвоста (джгутики) (рис. 9.1). Довжина їх в залежності від виду тварини варіює в межах 50 ... 80 мкм. Головка спермія становить приблизно 1/9 його довжини, має овальну форму, кілька усічену з боку шийки. В ході сперміогенеза з бульбашок комплексу Гольджі в цитоплазмі сперматіди формується акросома, яка являє собою обмежену мембраною орга Неллі, тісно прилягає до передньої частини ядра сперматозоїда і як шапочка одягаються його головку.
Акросома сперматозоїдів ссавців містить велику кількість вуглеводів, в ній присутні ферменти лізосом- ної природи (подібні до ферментами лізосом соматичних клітин) - гіалуронідаза, кисла фосфатаза і інші, а також протеолітичні ферменти (акрозін). З акросоми ферменти звільняються при контакті сперми з запліднює яйцем і, впливаючи на оболонки яйця, полегшують проникнення в нього сперми. З руйнуванням акросоми спермін втрачають здатність до запліднення, тому що не можуть проникати через оболонки яйцеклітини.
В ході сперміогенеза ядро сперматіди зменшується, набуває певну форму, в ньому відбувається конденсація хроматину, який перетворюється в щільний і метаболічно інертний. Мітохондрії шикуються в спіраль, обвиває хвіст сперматозоїда в його середній частині. Відбувається формування джгутика, що має складну будову. Основною структурою джгутика сперматозоїда є осьової комплекс, що складається з двох центральних мікротрубочок, оточених дев'ятьма периферичними парними мікротрубочками.
Сформовані спермін просуваються в просвіт канальця, де в слабощелочной середовищі набувають рухливість і надходять в прямі канальці і далі в звивистою канал придатка сім'яника.
Придаток сім'яника. Розташовується на поверхні насінники у вигляді тяжа. На придатку розрізняють головку, тіло і хвіст. Довжина каналу придатка сім'яника у самців коливається від 50 м (у баранів) до 85 м (у жеребців) і збільшується з віком тварини.
У стінках семявиносящіх канальців, що утворюють головку придатка, розрізняють епітеліальний і соедінітельнотканий шари. Епітеліальний шар складається з високих призматичних миготливих клітин, що містять ліпідні включення, і секреторних клітин, позбавлених війок. Ці клітини мають також здатністю до піноцитозу і фагоцитозу. Сім'явивідні канальці поступово зливаються в один більший канал, який утворює тіло і хвіст придатка. Просування сперміїв через придаток відбувається в основному за рахунок скорочення його м'язового апарату. Скорочення посилюються під дією окситоцину. У природних умовах посилене виділення окситоцину із задньої долі гіпофіза відбувається при статевому збудженні самця.
Під час проходження через придаток сім'яника спермии зазнають морфологічну і цитохімічних модифікацію. Вони набувають здатність до координації і до самостійного поступального руху, в них істотно зростає рівень АТФ. До мембрані спермія приєднуються вільні кислотні групи. Вони концентруються на хвості спермия, за рахунок чого тут утворюється підвищений негативний заряд, завдяки якому не відбувається аглютинація сперміїв. Таким чином, в процесі просування через придатки в сперміях відбуваються зміни, що підвищують їх стійкість до зовнішніх впливів і здатність до запліднення яйцеклітини. Густа мережа кровоносних судин і нервових волокон в стінках каналу придатка забезпечує необхідний склад середовища і швидке видалення з каналу продуктів обміну речовин.
Процес сперміогенеза займає тривалий час. Зокрема, у биків і баранів тривалість сперміогенеза і проходження сперміїв через канал придатка в середньому становить близько 50 діб. На проходження сперміїв через придаток сім'яника йде 6 ... 8 діб, отже, решту часу займає власне спермиогенез. У хвості придатка спермії можуть зберігати здатність до запліднення до 2 ... 3 міс.
Сім'япроводи і додаткові статеві залози. Сім'япроводи, куди далі надходять спермії, являють собою довгі тонкі трубки зі стінками з двох шарів гладкої мускулатури, вистелені циліндричним епітелієм. У бика і барана добре розвинені ампули семяпроводов і під час ерекції в них накопичуються спермії. У жеребця в ампулі семяпровода просвіт майже не розширений, а її стінки виконують роль секретирующие залози. Сім'япроводи відкриваються в тазову частину сечостатевого каналу. Сюди ж відкриваються протоки придаткових статевих залоз: пухирчастих, передміхурової, куперових і численних уретральних.
Парні пухирчастих залози знаходяться по боках від ампул семяпровода. У жеребця вони виробляють тягучий секрет, у бика, барана, кабана - рідкий. В їх секреті у великих кількостях містяться фруктоза, глюкоза, амінокислоти, інозитол, аскорбінова кислота, лужна фосфатаза. Секрет пухирчастих залоз стимулює метаболізм сперміїв.
Передміхурова залоза добре розвинена у жеребця і бика, слабо розвинена у барана, у кнура вона майже не розвинена. Секрет простати містить цитрат, аскорбінову кислоту, ліпіди, білки. Від секретів інших придаткових статевих залоз його відрізняє високий вміст цинку. Крім того, в секреті передміхурової залози у високих концентраціях присутні біологічно активні речовини - простагландини і вазогландін, що впливають на скоротливу здатність матки, а також протеолітичні ферменти. Куперови залози максимально розвинені у кнурів.
Відмінності в розвитку придаткових статевих залоз у різних видів тварин обумовлені типом осіменіння. У самок великої та дрібної рогатої худоби сперма при природному заплідненні, потрапляючи в піхву, пересувається потім в шийку матки, а у кобьш і свиней еякуліруєт безпосередньо в матку. Жеребці і кнури з розвиненими підрядними статевими залозами виділяють еякулят значно більший за обсягом, ніж бики і барани.
Значення секретів придаткових статевих залоз полягає в наступному: промивання і підготовка сечостатевого каналу до проходження сперми; збільшення обсягу еякуляту; стимулювання рухів сперміїв через сечостатевої канал; проштовхування сперміїв до вершини рогів матки (у свиней). Але, крім цих суто механічних функцій, безсумнівно, є ще й інші, недостатньо з'ясовані. Зокрема, в секреті передміхурової залози містяться такі біологічно активні речовини, як простагландини. Потрапляючи разом з еякулятом в статеві шляхи самки, вони підсилюють скорочення їх стінок, прискорюючи таким чином просування сперміїв по статевих шляхах самки.
Статеві рефлекси у самців. Функція розмноження у самців ссавців тварин являє собою складний комплекс рефлексів. Статевий потяг, або статевий інстинкт, виявляється в результаті сприйняття аналізаторами самця (слуховим, зоровим, нюховим, тактильним) подразнень, що виходять від самки, при достатньому рівні в крові андрогенів, перш за все тестостерону.
Роздратування від рецепторів надходять в кору великих півкуль, а звідти в гіпоталамус, що веде до посилення виділення гіпофізом гонадотропних гормонів, що збуджують діяльність сім'яників. Насінники, виділяючи в кров тестостерон, діють на центральну нервову систему, посилюючи збудження статевого центру та підтримуючи в ньому панівний осередок збудження - статеву домінанту. Порушення підкіркового статевого центру гормонами є силою, що діє на кору головного мозку і визначає поведінку тварини при проявах статевих функцій. Виконання статевих функцій відбувається при наявності умовних рефлексів, що виробляються на основі безумовних протягом життя тварини.
У самців сільськогосподарських тварин умовні статеві рефлекси виробляються надзвичайно швидко. Такий сильний подразник, як злучка або отримання сперми в штучну вагіну, веде до закріплення умовного статевого рефлексу з першого ж поєднання. Умовними статевими подразниками можуть бути вид случного манежу, подготовляемой до парування самки, надягання случной вуздечки і т. Д.
Крім позитивних рефлексів можуть вироблятися і негативні умовні рефлекси, які гальмують прояв статевої потенції: при різкому і грубому окрики, шумі, окрик, при неправильній підготовці штучної вагіни і при дії інших подразників, що заподіюють самцеві біль або викликають переляк в незнайомій обстановці.
Безумовні статеві рефлекси у самців є ланцюговими рефлексами, т. Е. Вони складаються з ряду взаємопов'язаних реакцій, причому завершення однієї з них є сигналом для виникнення наступної. У складному ланцюговому статевому рефлекс у самців розрізняють кілька основних стадій.