
| Головна |
| «« | ЗМІСТ | »» |
|---|
Основним катаболічсскім процесом деструкції глюкози в клітинах тварин і людини є послідовність ряду реакцій се окислення, в результаті яких в анаеробних умовах глюкоза перетворюється в лактат, а в аеробних - в кінцеві продукти: С02 і воду. Нижче наведена біологічна значущість окислювальних перетворень глюкози:
Гліколіз- це послідовність десяти ферментативних реакцій, в процесі яких в аеробних умовах глюкоза розщеплюється до двох молекул пірувату (аеробний гліколіз), а в анаеробних - до двох молекул лактату (анаеробний гліколіз). Нижче наведені стехиометричні рівняння процесів анаеробного (а) і аеробного (б) гліколізу:
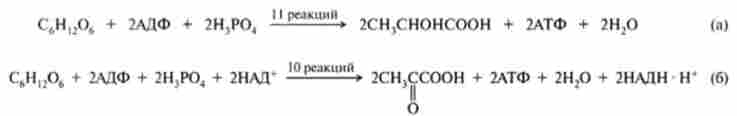
Поділ на анаеробний і аеробний гліколіз носить умовний характер, так як реакції гліколізу в присутності кисню і його відсутності одні й ті ж. Відмінності стосуються лише їх швидкості і кінцевих продуктів. При нестачі кисню реокісленіе НАДН, що утворився в ході гліколізу, здійснюється шляхом сполучення з відновленням пірувату в лактат, а в аеробних умовах НАДН окислюється в ході киснево процесу окисного фосфорилювання (гл. 15), результатом якого є утворення великої кількості АТФ.
Значення гліколізу. У клітинах значимість гліколізу полягає в наступному.
Послідовність реакцій гліколізу. Гліколіз протікає в дві стадії.
Процес гліколізу протікає в цитоплазмі клітини, каталізується одинадцятьма ферментами, більшість з яких виділено в високоочпшенном стані з різних джерел і добре вивчено.
Підготовча стадія гліколізу. У неї входять наступні реакції.
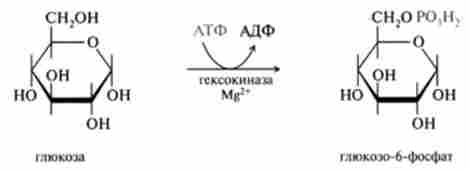
1. Необоротна реакція фосфорилювання глюкози і освіти глю- козо-6-фосфату, що каталізується ферментом гексокінази, описана раніше (18.2.1):
2. Оборотна реакція кето-альдольної ізомеризації глюкозо-6-фосфату у фруктозо-6-фосфат, що каталізується ферментом глюкозо-6-фосфатізоме- Разой:

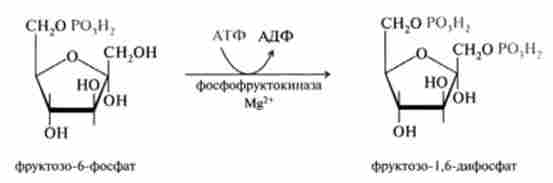
3. Необоротна реакція фосфорилювання фруктозо-6-фосфату молекулою АТФ до фруктозо-1,6-дифосфата, катализируемая ферментом фосфофрук- токіназой:
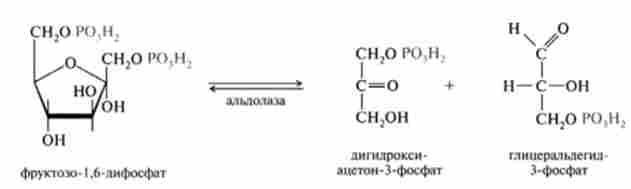
4. Оборотна реакція розщеплення зв'язку З-С у фруктозо-1,6-дифосфат на дві тріози, катализируемая ферментом альдолазой:
5. Оборотна реакція кето-альдольної ізомеризації дігідрооксіацетон- фосфату в глицеральдегид-3-фосфат, що каталізується ферментом тріозофос- фатнзомеразой:

Оскільки в наступні реакції включається тільки глицеральдегид-3- фосфат, у міру його споживання дегідроацетонфосфат перетворюється також в цю триоз. Таким чином, кожна молекула глюкози дає дві тріозние одиниці, які включаються в другу стадію гліколізу, тому необхідно, щоб кожна наступна реакція відбулася двічі.
Стадія генерації ЛТФ. У цю стадію входять наступні реакції.
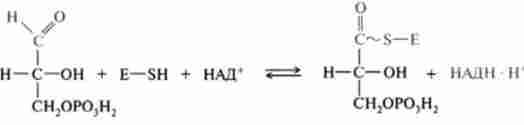
- фосфороліз утворилася тіоефірной зв'язку відбувається з приєднанням неорганічного фосфату, при цьому утворюється 1,3-діфосфогліце- рат і вільний фермент:
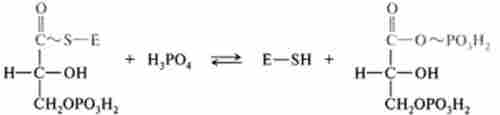
Сумарна реакція:
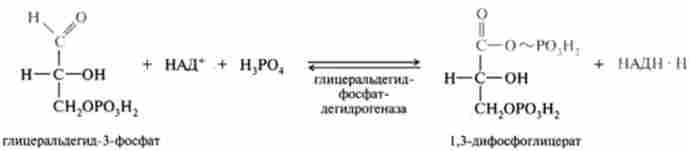
Таким чином, фосфороліз дозволяє потенційну енергію окислення альдегідної групи кумулировать в макроергічним фосфатного зв'язку 1,3-дифосфоглицерата в положенні 1. Реакцію окислення гліцер- альдегід-3-фосфату прийнято називати реакцією гликолитичної оксідоре- продукції.
7. Оборотна реакція субстратного фосфорилювання АДФ і освіти АТФ, при якій відбувається перенесення багатого енергією фосфорильної залишку з 1,3-дифосфоглицерата на АДФ. Реакція каталізується фосфогліце- раткіназой:
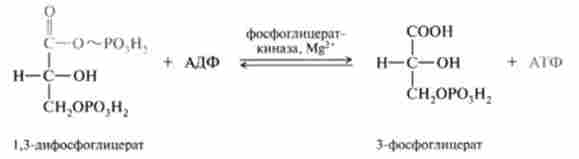
8. Оборотна реакція ізомеризації 3-фосфогліцерата в 2-фосфогліцерат, катализируемая ферментом фосфогліцератмутазой:

9. Оборотна реакція енолізаціі, в процесі якої відщеплення молекули води від 2-фосфогліцерата призводить до утворення макроергічних зв'язку в фосфосноілпіруватс. Реакція каталізується ферментом снолазой:
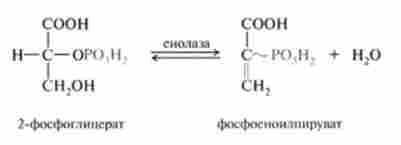
10. Ще одна реакція субстратного фосфорилювання АДФ і освіти АТФ, при якій відбувається розрив високоергічної зв'язку і перенесення фосфорильної залишку від фосфоеноілпірувата на АДФ. Каталізується ця реакція ферментом піруваткіназа; реакція практично необоротна:

11. Оборотна реакція відновлення пірувату до лактату (молочної кислоти) відбувається в анаеробних умовах за участю ферменту лактатде- гілрогенази і кофсрмснта НАДН - Н+:
Таким чином, в тканинах, що функціонують в умовах гіпоксії, спостерігається утворення лактату. Це особливо справедливо по відношенню до скелетного м'яза, інтенсивність роботи якої в певних межах не залежить від надходження кисню. Гліколіз в еритроцитах навіть в аеробних умовах завжди завершується утворенням лактату, оскільки в них відсутні мітохондрії, що містять ферменти аеробного окислення пірувату.
В цілому послідовність реакцій гліколізу представлена на рис. 18.4, молекулярні властивості гликолитичних ферментів наведені в табл. 18.1.
Таблиця 18.1. ферменти гліколізу
|
номер реакції |
фермент |
джерело |
Мовляв. маса. kDa |
кофермент, кофактор |
структура |
активатор |
інгібітор |
|
1 |
гексокіназа |
дріжджі |
96 |
Mg2* |
димер |
АДФ. Н3Р04 |
Глюкозо-6 фосфат. фосфосно- ілпірунат |
|
2 |
Глюкозо-6 фосфатізо- Мераз |
дріжджі |
145 |
Mg2' |
димер |
||
|
3 |
Фосфофрук- токіназа |
м'язи |
380 |
Mg2* |
тетрамер |
АДФ. АМФ. н3ро4, до * |
АТФ. цитрат. НАДН |
|
4 |
альдолаза |
м'язи |
160 |
тетрамер |
Zn *, Fe2*. з2* |
цистеїн |
|
|
5 |
Трнозофосфат- мзомераза |
Миіши |
56 |
Mg2' |
димер |
||
|
6 |
Гл і ис рал ьде- гід-3-фосфот- дегідро колагенази |
м'язи |
146 |
НАД * |
тетрамер |
Арсснат |
Mg2*, іодацетат |
|
7 |
Фосфоглі- іераткіназа |
Миіши |
50 |
Mg2* |
димер |
||
|
8 |
Фосфоглі- іератмутаза |
Миіши |
65 |
Mg2* |
димер |
||
|
9 |
Енолоза |
м'язи |
88 |
rMg2Mn ^ |
димер |
~ Слр |
|
|
10 |
Пмрунаткіна за |
м'язи |
240 |
K * |
тетрамер |
Mg2* |
Са ^ 'ТАТФ. ацетил-КоА. НАДН |
|
II |
Лактатдс- пшрогеназа |
м'язи |
240 |
НАД * |
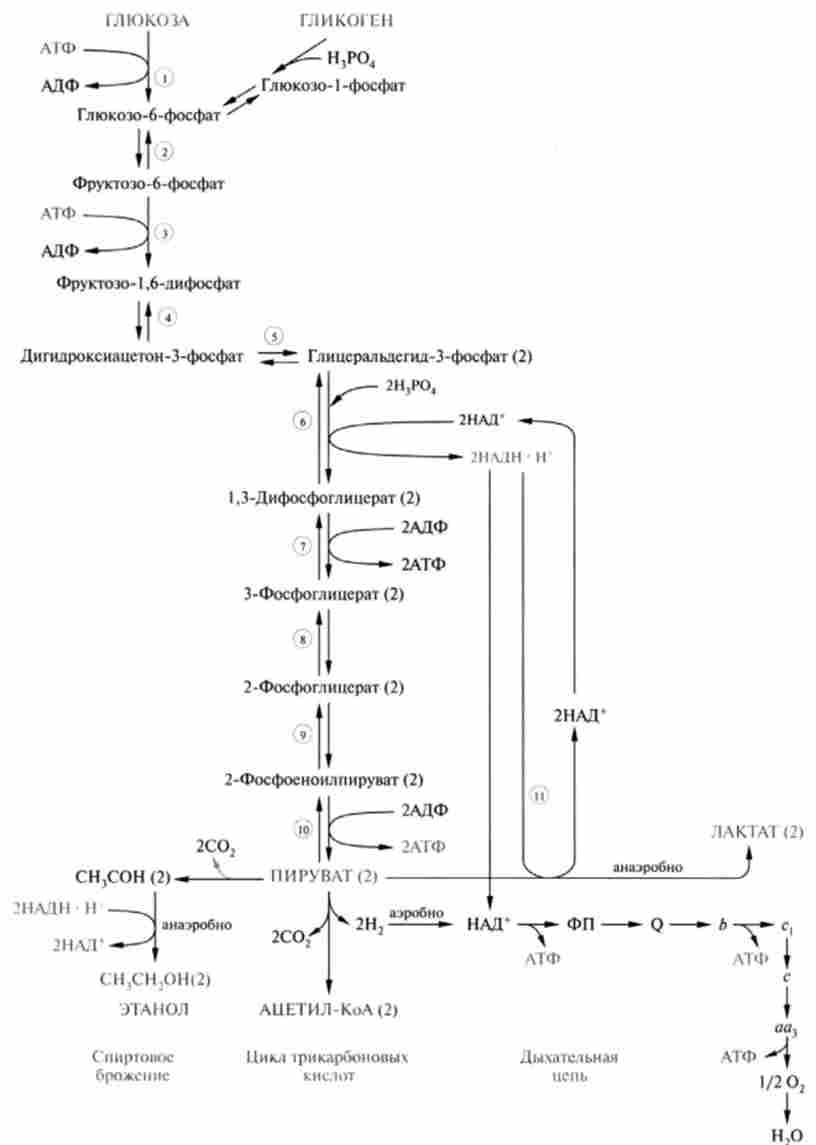
Мал. 18.4. Послідовність реакцій гліколізу, його зв'язок з аеробних окисленням глюкози, гликогенолизом, спиртовим бродінням: цифри в гуртку позначають номери реакцій: цифрою (2) - відзначені молекули, представлені двічі в розрахунку на одну молекулу глюкози