| Головна |
| «« | ЗМІСТ | »» |
|---|
Підшлункова залоза синтезує ферменти, необхідні для травлення. Ендокринна частина залози (острівці Лан- геранса) секретує принаймні чотири гормону: інсулін, глюкагон, сома- тостатін і панкреатичний поліпептид.
Структура і роль інсуліну були розглянуті вище. Глюкагон - 29-членний пептид, що викликає підвищення рівня глюкози в крові за рахунок стимуляції розщеплення глікогену в печінці, збільшує вміст глюкозо-6-фосфату в м'язах і володіє липолитичним (жиророзщеплювальних) дією.
Ці гормони вивільняються в панкреатичну вену, що впадає в ворітну вену печінки, що має важливе значення, оскільки для інсуліну і глюкаго- на печінку служить головною мішенню.
Основна роль інсуліну і глюкагону зводиться до регуляції вуглеводного обміну, однак гормони підшлункової залози впливають і на багато інших процесів. Наприклад, соматостатин пригнічує секрецію гормону росту. Він бере участь в локальній регуляції секреції інсуліну і глюкагону.
На рис. 12.15 приведена схема регулювання змісту глюкози СН в плазмі крові людини.
Відповідно до цієї схеми мозок (Сг), підшлункова залоза (Рп), печінку (Нр) та інші тканини (Т1) знаходяться в потоці крові, через який здійснюється гуморальна зв'язок між різними органами. Схема відображає одну зі специфічних особливостей взаємозв'язку метаболічних шляхів в організмі - просторове розділення взаємодіючих компонентів.
Більшість хімічних перетворень в організмі, а також активний транспорт через клітинні мембрани здійснюються за допомогою ферментів і білкових молекул (Е).
У схемі рис. 12.15 враховані тільки два гормону, що синтезуються в підшлунковій залозі, - інсулін (In) і глюкагон (G1).
Реакції в підшлунковій залозі:
Реакції в печінці:
Реакції в тканинах:
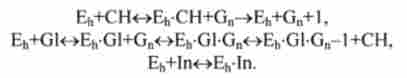
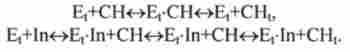
Реакції в крові (дезактивація гормонів):
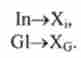
У наведених тут реакціях Е ,, Еь, Еа, Ер - ферменти, що функціонують у відповідних органах або тканинах; Рг - протеїн; In, G1 - гормони інсулін і глюкагон; СН, СН, - глюкоза в крові і тканинах, Gn - глікоген зі ступенем полімеризації і; Xj, Хс, - продукти дезактивації гормонів.
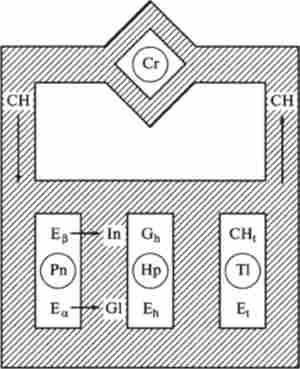
Мал. 12.15. Схема регулю вання концентрації глюкози СН в крові людини:
Сг - мозок. Рп - підшлункова залоза, Нр - печінку. TI - тканини. In - інсулін. GI - глюкагон. Єв. Е * - ферменти підшлункової залози, Еь - глікогснфосфорілаза, 1ц - фермент тканин, Оь - глікоген печінки.
СН, - глюкоза тканин
Роль інсуліну і глюкагону в життєдіяльності організму добре вивчена. Інсулін активує фермент Et і прискорює перехід глюкози в тканини скелетних м'язів і жирову тканину. В результаті такої активації рівень глюкози в крові швидко падає. Глюкагон активує фермент Еь - глікогенфосфорілазу - і прискорює процес деполімеризації полісахариду глікогену в печінці і м'язах з відщепленням глюкози. В результаті концентрація глюкози в крові зростає.
Збільшення вмісту глюкози в крові є стимулом для клітин підшлункової залози до збільшення активності ферменту Ер, що призводить до активації секреції інсуліну. Зменшення вмісту глюкози в крові призводить до активації ферменту Еа в а-клітинах цієї залози і збільшення секреції глюкагону.
На рис. 12.16 наведено типовий приклад автоколебательного характеру зменшення вмісту глюкози в крові людини після перорального введення різних доз цієї речовини.
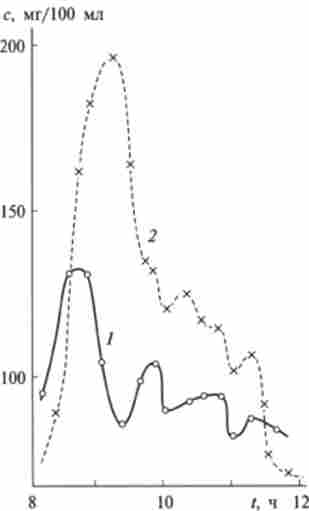
Мал. 12.16. Зміна концентрації глюкози з (Мг / 100 мл) від часу в крові здорової людини після перорального прийому всередину:
/ - 25 і 2 100 г глюкози (норма 70-110 мг / 100 мл (4-6 мМ))
Схему регулювання рівня глюкози в крові неважко описати за допомогою кінетичних рівнянь. Аналіз цих рівнянь дозволяє зробити висновок, що автоколебательний характер повернення біохімічного складу організму до вихідного стану після імпульсного збурення пов'язаний з наявністю принаймні однієї автокаталітічсской реакції в метаболічному шляху. Такі реакції є в розглянутої тут схемою реакцій регулювання рівня цукру в крові.
Наприклад, збільшення концентрації глюкози в відповідно до реакції Ер+ СН Ер-СН -> Ер + In призводить до збільшення вмісту інсуліну в крові,
а це викликає збільшення абсорбції глюкози тканинами м'язів
(Е, + In Et-In + СН E, -In + СН E, In + СН,).
Крім автоколебаний слід звернути увагу на інші особливості регулювання цукру, які ілюструє рис. 12.16.
По-перше, максимальна кількість глюкози в крові зростає непропорційно дозі: дози розрізняються в 4 рази, максимальні кількості - приблизно лише в 1,5 рази. Ці максимальні значення в кілька разів нижче гранично можливих при даних дозах 25 і 100 г. Максимальне значення при меншій дозі досягається значно швидше, ніж при великій. Крім того, спостерігається «проскакування» початкової кількості глюкози в крові до кінця періоду релаксації до стаціонарного значення.
Перераховані особливості адаптаційного регулювання в тій чи іншій мірі проявляються і для інших біогенних речовин, що вводяться в організм. Вони пов'язані з характером всмоктування або введення в кров, з одного боку, і характером реакційної ланцюга перетворень - з іншого. Практична відсутність періоду затримки вказує на швидке включення компенсаційного механізму адаптації внутрішнього середовища.
Експериментальне дослідження регулювання концентрацій іонів ОН, CI, амінокислот і інших речовин в організмі показує, що регулювання і цих компонентів носить автоколебательний характер. Це загальна властивість процесів регулювання рівня різних речовин в крові, очевидно, відображає спільність механізмів регулювання, в основі яких лежать реакційні кола типу схеми регулювання рівня глюкози.
Наведені вище факти дозволяють зробити висновок, що регулювання температури тіла також має демонструвати автоколебательний характер досягнення рівноважного значення. Цей висновок підтверджується експериментальними даними.
Зміна температури в організмі гомойотермних (теплокровних) тварин викликає відповідну реакцію терморецепторов. Охолодження тканин призводить до збільшення витрати енергії для підтримки температури на необхідному рівні, що, в свою чергу, інтенсифікує процеси поглинання глюкози тканинами і відповідне зменшення концентрації глюкози в крові. В результаті включається механізм компенсації, що відображається реакціями регулювання рівня глюкози.