| Головна |
| «« | ЗМІСТ | »» |
|---|
Таким чином, в момент нанесення на возбудимую клітку зовнішнього подразника в неї протягом дуже короткого часу входять іони натрію, і мембрана клітини деполяризуется настільки, що спостерігається овершут, але в наступну мить різко зростає струм До+ з клітки, в результаті чого потенціал на мембрані повертається до рівня спокою: -70 мВ. Нагадаємо, що в спокої мембрана збудливих клітин цілком проникна для К+, а для Na+ є практично непереборною перешкодою.
Так яким же чином забезпечуються струми іонів через мембрану в момент генерації ПД? Виявляється, в клітинній мембрані збудливих клітин розташовуються особливі білкові молекули, що утворюють іонні канали. Висхідна фаза ПД реалізується за рахунок роботи потенціалзавісімих натрієвих каналів. Коли клітина знаходиться в стані спокою, цей канал закритий (рис. 8.8).
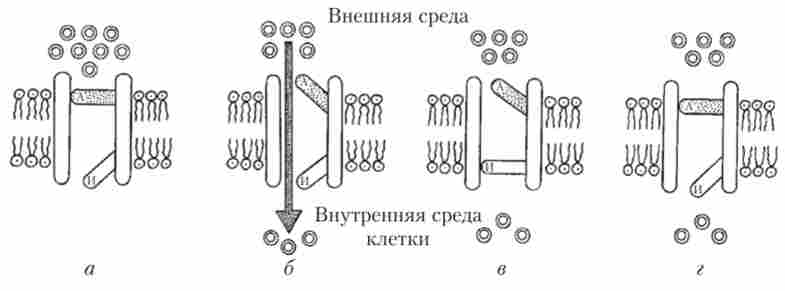
Мал. 8.8. Схема роботи потенціалзалежне натрієвого каналу:
а - канал закритий; 6 - канал відкритий; в - канал инактивирован; г - канал повернутий в початковий стан і готовий до роботи; А - активаційна стулка;
І - інактіваціонная стулка
Якщо ж клітку дратують будь-яким зовнішнім стимулом, то ПП на мембрані починає зрушуватися від -70 мВ до -50 мВ. Для більшості нервових клітин досить зсуву ПП на 20 мВ (т. Е. До -50 мВ) для того, щоб просторова конфігурація білкової молекули каналу змінилася, утворюючи прохід, по якому в клітку спрямовуються за градієнтом концентрації іони натрію. Відбувається це через те, що частина молекули каналу (яку назвали активационной стулкою каналу) зсувається, і відкривається канал для іонів натрію. Однак наступної миті інша частина канальної молекули білка (названа інактіваціонной стулкою каналу) перекриває рух іонів через канал (див. Рис. 8.8).
Натрієвий канал знаходиться у відкритому стані всього 1 мс, але за цей час через нього в клітку встигає пройти близько 6 тис. Іонів Na+. Звичайно, робота одного каналу не може помітно деполярізовать клітку, але натрієвих каналів на мембрані збудливих клітин дуже багато - близько 100 на площі в 1 мкм2. Після спрацьовування канал за кілька мілісекунд
повертається в початковий стан і може відкритися знову, якщо клітина піддасться подразнення (див. рис. 8.8). Але для того, щоб Na+-Kana.T перейшов зі стану, зображеного на рис. 8.8, в, в стан, зображене на рис. 8.8, а, необхідно, щоб потенціал на мембрані досяг ПП. Відновлення ДГП на мембрані здійснюється завдяки тому, що відразу ж після закриття натрієвого каналу через свої особливі канали з клітки виходять позитивно заряджені іони К+. Калієві канали діють за тими ж принципами, що і натрієві, однак у них є тільки одна стулка, яка відкриває і закриває шлях іонів калію з клітки.
Іонні канали мають вибірковістю: пропускають тільки «свої» іони. Однак бувають і винятки. Наприклад, іони літію (Li+) Дуже добре проходять через натрієві канали всередину збудливих клітин (навіть трохи краще, ніж сам натрій). Але літію в організмі людини практично немає. Літій застосовується для зміни роботи мозку в разі деяких важких психічних розладів.
В даний час відкрито кілька видів натрієвих і калієвих каналів. Вони розрізняються величиною порога відкривання, швидкістю відкривання і закривання, пропускною спроможністю для іонів, механізмом відкривання.
Існують також особливі малоізбірательние канали для аніонів та катіонів і, навпаки, дуже виборчі канали для Са2+ і С1 ".
Відразу після того, як активаційна стулка пропускає в клітку порцію натрію, канал знову активувати неможливо (див. Рис. 8.8), так як необхідно спочатку повернути в початковий стан інактіваціонную стулку. У цьому стані канал називають абсолютно рефрактерним, т. Е. Нездатним до роботи. За тривалістю абсолютна рефрактерність дорівнює приблизно 1 мс. Наявність абсолютної рефрактерності не дозволяє окремим ПД накладатися один на одного і обмежує максимальну частоту розрядів нервових клітин величиною 500-700 Гц. Слідом за стадією абсолютної рефрактерності настає відносна рефрактерність: лише частина Иа+-Капали повернулася в стан готовності, ще не закриті всі потенціалзалежні До+-канали. В таких умовах запуск ПД можливий, але тільки під дією більш потужного стимулу.
Нормальна робота іонних каналів дуже важлива для організму. Адже порушення їх роботи призводить до дуже швидкого розвитку порушень функціонування ЦНС, судом, паралічів і т. І.- в залежності від того, який вид каналів дезорганізований.
Природа дуже далеко зайшла по шляху створення блокаторів іонних каналів, якими по справедливості озброїла і хижаків, і тих, хто від цих хижаків обороняється.
Один з найсильніших отрут міститься в тканинах риби-іглобрюха, званої японцями рибою фугу. У цих тропічних риб особливо отруйні печінку і яєчники. Незважаючи на це, іглобрюх є делікатесом японської кухні. Отрута з тканин цих риб називається тетродотоксином, і досить потрапляння в організм людини всього лише 0,5 мг токсину, щоб він загинув. Для порівняння: знаменитий ціаністий калій приблизно в 100 разів менше отруйний. Тетродотоксин діє з зовнішньої сторони натрієвого каналу, блокуючи вхід в нього натрію. В результаті генерація ПД стає неможливою, що призводить до розслаблення мускулатури і паралічу дихання.
Ще більш страшна отрута міститься в шкірі зовні абсолютно нешкідливих жаб з роду листолазів, що живуть в Південній Америці. Ця отрута, званий батрахотоксин, ядовитее ціаністого калію в 500 разів! Одна маленька жаба носить в шкірі стільки отрути, що їм можна погубити до 350 осіб. Батрахотоксин блокує натрієвий канал в момент його відкриття, не даючи стулок зачинитися, після чого він не може закритися, і натрій, вступаючи в клітку без обмежень, призводить до її надмірного збудження. В результаті, як правило, першими нс витримують клітини серцевого м'яза, і серце перестає битися.
Схожим ефектом володіють отрути скорпіонів, що підтримують натрієві канали у відкритому положенні. Блокатори натрієвих каналів виробляють і водорості (САК- сітоксін), і вищі рослини (аконітін).
Ще одна група речовин, що надають свій лікувальний ефект через вплив на натрієві канали, - місцеві анестетики (новокаїн, лідокаїн, прокаїн і багато інших). Молекули цих речовин проникають в канал зсередини клітини, знижуючи натрієві струми і блокуючи проведення електричних сигналів але тонким безмякотним нервових волокнах. Якраз за такими волокнам передасться в мозок інформація про больових впливах. Таким чином, місцеві анестетики на час блокують больові відчуття.
блокаторів К+-каналів в природі мало, зате створений синтетичний блокатор - тетраетіламмоній, «затикає» канал зсередини клітини.
В даний час відомо безліч типів Na+- і К+-каналів, характерних для тих чи інших клітин організму. Ці канали відрізняються один від одного умовами, в яких вони відкриваються і закриваються, пропускною спроможністю, швидкістю відкривання і закривання і т. П. Крім того, знайдені канали для С1 ~, Са2+ і т.п.
Значну роль у всіх збудливих тканинах відіграють кальцієві струми, що йдуть по градієнту концентрації в клітину. Існують специфічні потенціалзалежні канали, через які Са2+ входить в клітку, виконуючи при цьому дві функції: як позитивно заряджений іон - деполяризує мембрану і як потужний внутрішньоклітинний регуляторний фактор - змінює роботу всіх систем клітини.
Дуже важливі кальцієві струми в процесах збудження м'язів і передачі інформації від клітини до клітини. В еволюції канали для кальцію, мабуть, виникли дуже рано і Са2+-струм через мембрану древнє Ка+-струму. Канали для кальцію відкриваються відносно повільно, щільність їх на мембрані менше, ніж каналів для Na+, але зате Са2+-канали не закриваються до кінця, забезпечуючи постійний слабкий вхід цього іона в клітку. Блокаторами кальцієвого струму через канали є багато двовалентні іони: кадмій, кобальт, марганець і ін.
На закінчення цього параграфа ще раз порівняємо роботу іонних каналів і іонних насосів (Иа+/ К+-АТФ-аза). Завдання обох систем - транспорт іонів через мембрану збудливою клітини, але на цьому їх схожість і закінчується.
Порівняння пасивного (канали) і активного (насоси) транспорту Na+ і К+ через клітинну мембрану
Таблиця 8.1
|
характеристика |
канали |
насос |
|
Напрямок руху іонів |
За електрохімічного градієнту |
проти градієнта |
|
Джерело енергії |
градієнт іонів |
енергія АТФ |
|
характеристика |
канали |
насос |
|
Залежність провідності від потенціалу на мембрані |
залежить |
Не залежить |
|
блокатори транспорту |
тетродотоксин |
Дигітоніну, сапонін, строфантин |
|
вплив температури |
вплив мале |
Охолодження гальмує транспорт |
|
швидкість транспорту |
Під час висхідної фази ПД - 10 s моль / см2 - з |
6 - 10 "11 моль / см2 - з |
|
Вплив блокаторів енергетичного обміну |
Чи не впливають |
Блокатори припиняють роботу насоса (ціаніди, динитрофенол) |
Очевидно, що транспорт іонів через канали і насоси - абсолютно різні незалежні процеси, проте активний транспорт (насоси) абсолютно необхідний для того, щоб забезпечити підтримку різниці змісту Na+ і К+ по різні боки мембрани, що і визначає можливість збудливою клітини утримувати ПП і генерувати ПД шляхом переміщення іонів через канали.