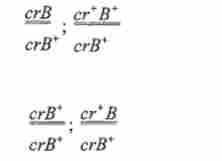
| Головна |
| «« | ЗМІСТ | »» |
|---|
Рекомбінація генів здійснюється різними способами. Цей процес може бути пов'язаний з перерозподілом цілих хромосом. Такий механізм відповідно до третього закону Г. Менделя забезпечує незалежне успадкування незчеплених генів і ознак. Найчастіше рекомбінацію у вузькому сенсі слова пов'язують з кросинговером, т. Е. З пе- рекомбинацией генів, локалізованих в гомологічних хромосомах. Цей тип рекомбінації розглядається в цій главі.
У профазі I мейозу на стадії діплотени у багатьох організмів добре помітні характерні фігури, утворені гомологічними хромосомами, - хіазми. У 1909 р Ф. Янссену припустив, чгго освіту хиазм пов'язано з обмінами гомологічними ділянками гомологічних хромосом. Пізніше Т. Х. Морган зв'язуй хіазми з кросинговером, що відбувається відповідно до гіпотези К. Бріджеса (1915) по механізму розрив-возз'єднання.
С. Дарлінгтон, виходячи з числа хиазм в пахітене, визначив загальну довжину кожної з хромосом кукурудзи. При цьому він прийняв, що одна хіазма утворюється на ділянці в 50 сМ. Отриманий на підставі цитологічних даних результат добре узгоджувався з довжинами груп зчеплення, обчисленими звичайним способом виходячи з частот кросинговеру при гибридологічний аналізі. Паралелізм подій, що реєструються генетично і цитологічних при рекомбінації зчеплених генів, був продемонстрований в 1931 р для двох об'єктів: кукурудзи і дрозофіли. В обох випадках був використаний принцип: зіставлення результатів кросинговеру з фізичними обмінами гомологічних ділянок гомологічних хромосом. Очевидно, для такої роботи схрещувати форми повинні бути дігетеро- зиготности як за генетичними, так і по цитологічним маркерами однієї пари хромосом.
Харрієт Крейтон і Барбара МакКлінток вирішили цю проблему щодо кукурудзи. Була досліджена форма, дігетерозіготная по генам: з (colorless) - безбарвний алероновий шар ендосперму, рецесія; з+ - пофарбований, домінант; wx (yvaxy) - ендосперм, а також пилкові зерна, які містять амилопектин, внаслідок чого фарбуються йодом в червоний колір, рецесія; wx - ендосперм і пилкові зерна, які містять амилозу, фарбується йодом в синій колір, домінант.
Крім того, досліджена форма на одному кінці хромосоми несла блок (ділянка) гетерохроматина, а на іншому - транспонований ділянку іншої хромосоми. Другий гомолог цих морфологічних відмінностей не мав.
У потомстві відбирали два реципрокних класу, рекомбінантних по з і wx.
Рослини, вирощені з насіння цих рекомбинантов, досліджували цитологічних і переконалися в тому, що у всіх рекомбинантов за генетичними маркерами стався фізичний обмін ділянками гомологічних хромосом.
Подібним чином вирішив ту ж проблему К. Штерн для дрозофіли. Він досліджував розщеплення в потомстві від схрещування самок, дігетеро- зиготних по генам Л'-хромосоми: cr (carnation) - очі червоні, рецесія; сг - нормальна темно-червоне забарвлення очей, домінант; В {Ваг) - полосковідние очі, домінант: В * - круглі очі, рецесія (рис. 6.1).
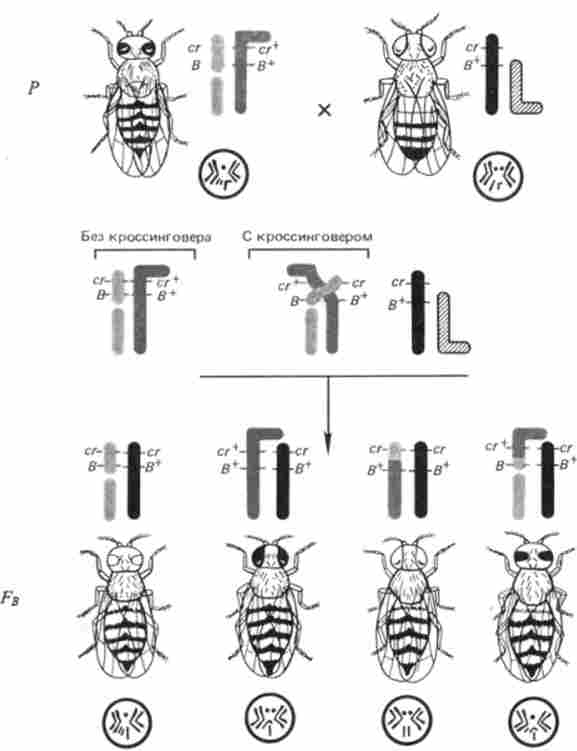
Мал. 6.1. Схема експерименту К. Штерна, який демонструє відповідність між рекомбинацией генів і фізичними обмінами ділянками Л'-хромосоми у дрозофіли
Ці ж самки несли гетероморфними Х-хромосоми: одну сильно вкорочений внаслідок транслокації її бесцентромерного ділянки на четверту мікрохромосому, іншу - Г-подібну внаслідок транслокації на неї фрагмента У-хромосоми. Таких самок схрещували з самцями, що несуть рецесивні аллели досліджуваних генів (сг, В+) В Х-хромосомі звичайної форми і мали звичайну У-хромосому.
Результат кросинговеру у самок в такому схрещуванні - рекомбінантні класи - вивчали цитологічних. Для цієї мети брали лише самок, оскільки самці мають субметацентрічні, т. Е. Разноплечую У-хромосому, яку можна сплутати з Г-образної ^ хромосомою.
Всі чотири класи самок-нащадків відрізнялися один від одного за фенотипом:
1) нерекомбінантні:
2) рекомбінантні:
а також за структурою своїх X-хромосом.
Дослідивши 374 самки, К. Штерн показав, що кросинговер між генами сг і В супроводжувався фізичним обміном між Л'-хромосомами.
Експерименти X. Крейтон і Б. МакКлінток з кукурудзою, К. Штерна з дрозофіли довели, що в основі кросинговеру лежить реальний обмін ділянками гомологічних хромосом, однак механізм цього обміну залишився незрозумілим. Поряд з гіпотезою К. Бріджеса (механізм розрив-злиття) розглядався і інший механізм, запропонований Дж. Беллінгем (1933), вивчав мейоз у рослин. Відповідно до цієї гіпотези, розривів і перевоссоедіненія хромосом не відбувається, а кросинговер приурочений до стадії відтворення хромосом. При цьому, як вважав Дж. Беллінг, спочатку відтворюються хромомери, потім їх з'єднують хромонеми. Таке з'єднання може привести до рекомбінантним сполученням хромомер. У 1930 р X. Винклером була запропонована ще одна гіпотеза, згідно з якою в потомстві дигетерозиготи рекомбінантні класи можуть з'являтися внаслідок спрямованих змін алелей під впливом один одного - гіпотеза конверсії:
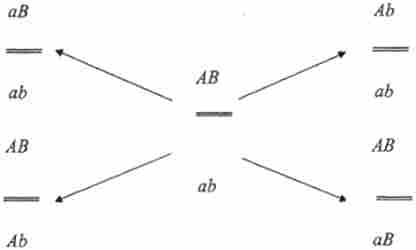
Зошитовий аналіз у грибів показав, що подібні зміни як правило не відбуваються. Чи не узгоджувалися з цією гіпотезою і дані цитологічного вивчення результатів кросинговеру. Однак остаточно з'ясований механізм реципрокного обмінів і дознавств фізичний обмін ділянками батьківських молекул ДНК при рекомбінації значно пізніше - в 1961 р в експерименті М. Мезельсоном і Дж. Уейгла, які вивчали рекомбінацію у бактеріофага X.
В експерименті використовували біологічні (генетичні) і фізичні (ізотопи ,4З і 1SN) маркери, за якими контролювали фізичні обміни ділянками генетичного матеріалу при рекомбінації.
Доказ справедливості гіпотези розрив-возз'єднання було отримано також Дж. Тейлором в 1967 р для коника.