| Головна |
| «« | ЗМІСТ | »» |
|---|
збудливість - здатність клітин і тканин переходити в стан фізіологічної активності йод впливом будь-яких зовнішніх впливів (подразників), які досягли порогової величини. Збудливістю мають будь-які живі клітини, в тому числі рослинні, але найбільшою мірою це знаходить свій вираз у клітин тварин - таких, як нервові, м'язові і залізисті.
Будь-якому фахівцеві, пов'язаному з дослідженнями організму тварини і людини, необхідно розуміти, що саме збудливі клітини складають основу НС, сприймають сигнали із зовнішнього та внутрішнього середовища організму, забезпечують його відповідні реакції.
Все збудливі клітини володіють трьома властивостями:
1) ПН, або мембранним потенціалом, - різницею електричного потенціалу між внутрішньоклітинної середовищем і середовищем, що оточує клітину;
Проведемо наступний експеримент. Якщо взяти нейрон, що знаходиться в даний момент в стані спокою, і ввести тонкий скляний мікроелектрод з кінчиком діаметром не більше 1 мкм, то такий кінчик не нанесе клітці видимих пошкоджень. Порожнина скляного електрода повинна бути заповнена рідиною, добре проводить струм (електролітом). Найчастіше для цієї мети використовують розчин хлористого калію (КС1). Електрод з'єднують з вольтметром. Поки кінчик електрода знаходиться в міжклітинному середовищі, стрілка мікровольтметр стоїть на нулі (рис. 8.1).
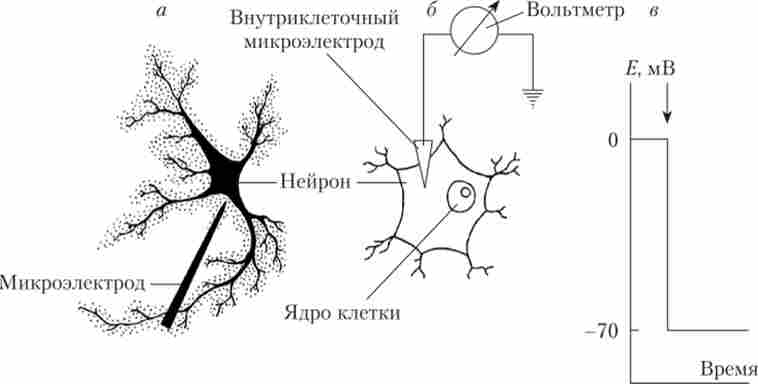
Мал. 8.1. Реєстрація потенціалу спокою нейрона:
а - малюнок з мікрофотографії; б - схема реєстрації потенціалу спокою шляхом
введення в нейрон кінчика мікроелектрода; в - стрибок мембранного потенціалу в момент введення в нейрон кінчика мікроелектрода (4)
У момент проколювання мембрани нейрона реєструється стрибок потенціалу вниз до рівня приблизно -70 мВ. Це і є мембранний потенціал, або ПП. Якщо електрод не рухаються, а для нейрона створити правильні умови (склад навколишнього розчину, температура), то ПП буде підтримуватися без всяких видимих змін протягом декількох годин. ПП знайдений у всіх збудливих клітинах, і його величина коливається від -30 до -100 мВ в залежності від того, з якими клітинами проводиться досвід.
Потенціал спокою був відкритий в середині XIX в. великим швейцарським фізіологом Емілем Дюбуа-Реймон а його учень Юліус Бернштейн створив найпершу теорію, яка пояснювала, чому всередині збудливих клітин, що знаходяться в стані спокою, реєструється явний надлишок негативних зарядів. Виходячи з даних, отриманих цілою низкою дослідників, Бернштейн частково встановив, а частково припустив наступне:
Якщо це так, то частина іонів К+ повинна виходити через клітинну мембрану в міжклітинну середу, залишаючи в клітці надлишок парних До+ негативно заряджених частинок - аніонів.
Тому на внутрішній поверхні мембрани клітини створюється негативний заряд (рис. 8.2).
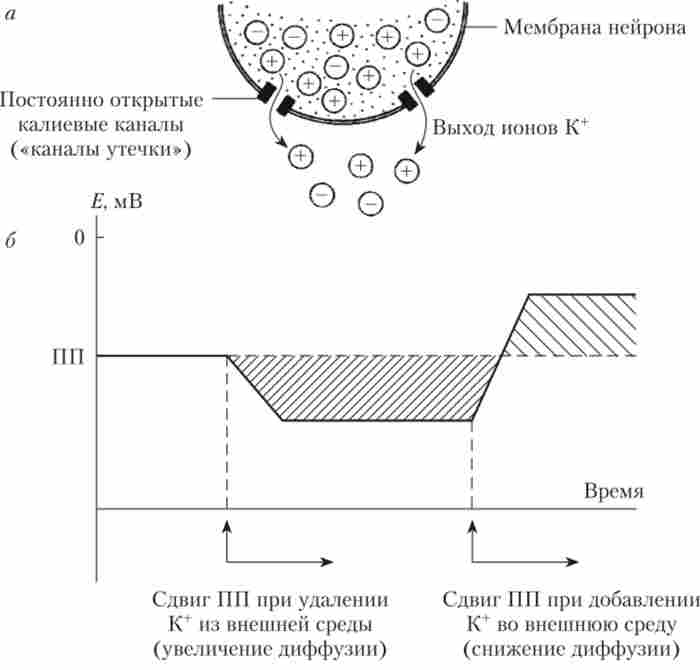
Мал. 8.2. Роль іонів К+ у виникненні і підтримці потенціалу спокою (ПП):
а - виникнення ПП внаслідок дифузії іонів К+ через постійно відкриті канали витоку; б - зміна рівня ПП при зміні концентрації К * у
зовнішньому середовищі
Але надлишок негативно заряджених частинок буде притягувати до себе іони К+, перешкоджаючи їх виходу з клітини, а вийшли в міжклітинний простір іони К 'також будуть «відштовхувати» позитивно заряджені частинки, не даючи вийти з клітки новим порцій До+. У підсумку вихід К+ триває лише до тих пір, поки сила дифузії (концентраційного тиску) і сила електричного нуля не стануть рівні. Точка рівноваги і відповідає рівню потенціалу спокою.
Струм іонів при досягненні ПП не зупиняється, так як є постійно відкриті канали і К+ продовжує переміщатися крізь мембрану, але число іонів, які увійшли в клітку і вийшли з неї, тепер виявляється однаковим. Такий стан називається динамічною рівновагою - рівністю двох протилежно спрямованих процесів. Якщо один з процесів посилиться або ослабне, то точка рівноваги зміститься. Наприклад, якщо, як це робив Бернштейн, штучно збільшити концентрацію К+ в міжклітинному середовищі, то це, природно, буде заважати виходу з клітини нових порцій позитивно заряджених частинок (К+), І ПП стане менш негативним, зміщуючись до нуля (див. Рис. 8.2). Якщо ж штучно зрівняти концентрацію К+ в цитоплазмі і поза клітиною, то ПП дорівнюватиме нулю. Так Бернштейн довів, що ПП в збудливою клітці визначається різницею концентрації К+ всередині цієї клітини і в позаклітинному середовищі. Бернштейн запропонував для кількісної оцінки ПП використовувати рівняння, виведене Вальтером Нернстом для штучної системи, яка представляє собою два розчину КС1 різної концентрації, розділених штучної мембраною, проникною тільки для До+ (Рис. 8.3).
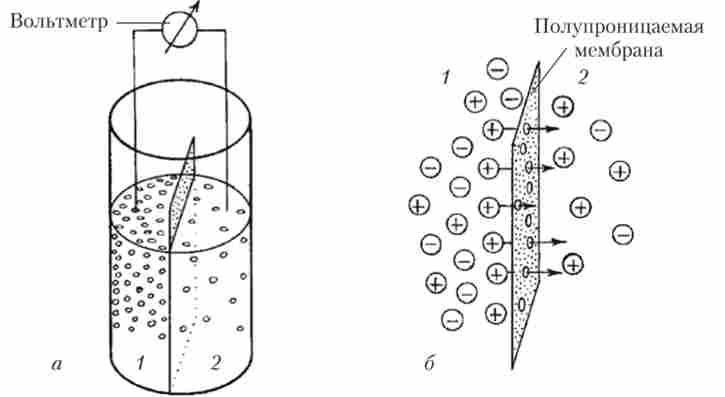
Мал. 83. Схема досвіду Вальтера Нернста:
а - принципова схема установки з двома судинами (1 і 2), що містять різні концентрації КС1 і розділеними мембраною, проникною тільки для До+;
6 - потік (показаний стрілками) катіонів (+) через мембрану з області з їх більшою концентрацією (1) в область з меншою концентрацією (2), що приводить до виникнення на мембрані потенціалу
В одну посудину він налив 10% -ний розчин КС1, а в другій - 1% -ний розчин цієї солі. В обох розчинах сталася дисоціація КС1 на До+ і СП, але в посудині 1 початково було в 10 разів більше і катіонів (К+), І аніонів (С1 "), ніж в посудині 2. Так як розділяє розчини напівпроникна мембрана добре пропускає катіони, то частина іонів калію (К+) Перейшла з посудини 1, де концентрація КС1 початково була вище, в посудину 2, в якому концентрація КС1 була в 10 разів нижче. Оскільки іони калію несуть позитивний заряд, то позитивних зарядів в посудині 2 буде більше, ніж негативних. У посудині 1 таким чином виявиться деякий надлишок «кинутих» аніонів хлору, які втратили свої катіони калію. Але негативно заряджені іони С1 "будуть притягувати назад частину позитивно заряджених іонів К+ за рахунок взаємодії електричних зарядів. Через якусь мить потоки До+ з посудини 1 в посудину 2 і навпаки стануть рівними. Однак в посудині 1 іонів К+ буде все одно менше, ніж іонів С1_ (Адже частина К * перейшла в посудину 2), а в посудині 2 буде більше іонів К+, ніж іонів С1_ (За рахунок тих, хто прийшов з посудини 1). Отже, розчин в посудині 1 буде заряджений негативно по відношенню до розчину в посудині 2.
Між розчинами встановиться нернстовскую потенціал, який, відповідно до рівняння Нернста, дорівнює
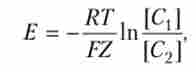
де Е - величина потенціалу, що виникає між розчинами, налитими в посудину 1 і посудину 2; R - газова постійна; Т - температура; F - число Фарадея; Z - валентність; [З{ - концентрація іонів К+ в посудині 1; [З2] Концентрація іонів К+ в посудині 2.
Бернштейн запропонував використовувати наведене вище рівняння Нернста для визначення мембранного потенціалу, як тільки дізнався, що саме До+ може проходити через мембрану спочиває збудливою клітини. Однак співвідношення концентрацій катіонів калію всередині і зовні клітини виявилося не 1: 10 (як в штучної експериментальної системі Нернста), а зовсім іншим. Так, в м'язових клітинах калію в 49 разів більше, ніж в середовищі, що оточує клітини. Зате в навколишньому середовищі, т. Е. Навколо збудливих клітин, іонів натрію (Na+) Приблизно в 10 разів більше, ніж усередині клітин. Однак, коли клітина знаходиться в стані спокою, натрій не може проникати крізь клітинну мембрану всередину клітини і не бере участі в створенні 1111. За останні 70 років багато вчених шляхом оригінальних експериментів остаточно довели, що негативний заряд спочиває збудливою клітини обумовлений тим, що частина іонів калію виходить через зовнішню мембрану в зовнішнє середовище до тих пір, поки не встановиться динамічна рівновага: скільки іонів калію вийшло з клітки, стільки ж і ввійде назад. При цьому на мембрані встановлюється ПП, характерний для даного виду збудливих клітин і визначається тим, що в цитоплазмі клітини залишається надлишок негативно заряджених іонів - аніонів, головним чином, великих білкових молекул, що мають негативний заряд, «покинутих» катіонами, т. Е. Іонами калію. Аніони пройти через мембрану не можуть і залишаються в клітці, визначаючи негативний заряд цитоплазми.
Однак згодом було показано, що в мембрані нейронів є невелика кількість постійно відкритих каналів для калію, за якими катіони натрію за градієнтом концентрації (навколо нейрона в 10 разів більше натрію, ніж в цитоплазмі) постійно проникають у клітину, зрушуючи 1111 до менш від'ємних показників.
Вхідний в клітку невеликий струм натрію (струм витоку), змінюючи IIII, робить нейрон більш збудливим; ніж цей струм значніше, тим сильніше IIII зрушать до тих пороговим величинам, за якими слід перехід нейрона в стан збудження. Таким чином, виникає можливість підтримувати нейрони одних систем на високому рівні збудливості, а інших - на зниженому. Як було з'ясовано, в міжклітинному середовищі міститься в п'ять разів більше іонів хлору (С1 "), ніж в цитоплазмі, проте в спокої ток іонів хлору через зовнішню мембрану в клітину дуже малий і становить всього 4%, якщо порівняти з струмом іонів калію, але , строго кажучи, для точного визначення величини ДГП в рівнянні Нернста треба підставити величини проникності для всіх видів іонів, які можуть в спокої проникати через мембрану клітини і міняти ПП. Отримане рівняння носить ім'я Голдмана - Ходжкіна - Катца:
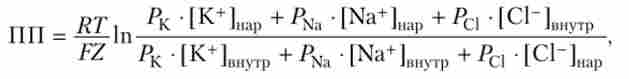
де Р - проникність мембрани для відповідних іонів.
Механізм генерації ПП, концентрація іонів поза і всередині клітин, проникність зовнішньої мембрани для іонів - все це досліджувалося на різних об'єктах, однак основний вид клітин, на якому проводилися досліди, - нейрони кальмарів (точніше, аксони цих нейронів). У 1930-і рр. було виявлено, що аксони нейронів деяких нервових вузлів (зірчастого ганглія) цих головоногих мають гігантський діаметр (до 1 мм), і з ним дуже зручно експериментувати.
Отже, ПП збудливою клітини визначається декількома факторами, головними з яких, безумовно, є різниця концентрацій К+ всередині і зовні клітини і вільний струм До+, що йде через мембрану. Певну роль в генерації ПП грають слабкі входять струми Na+ і С1 ", для яких мембрана клітини, що знаходиться в стані спокою, майже непроникна. Змінюючи будь-який з цих параметрів, можна в тій чи іншій мірі змінювати ПП, тим самим змінюючи здатність клітини до порушення.