| Головна |
| «« | ЗМІСТ | »» |
|---|
Мейоз - це два наступних один за одним поділу клітини, які лежать в основі утворення гамет, що містять один набір (П) хромосом, на відміну від соматичних клітин, що мають два набори (2л) хромосом.
Повною мірою це справедливо для гаметогенеза тварин, т. К. Під час мейозу у них утворюються гаплоїдні клітини, що диференціюються і функціонують як гамети: яйцеклітини і сперматозоїди. У вищих рослин внаслідок мейотічних поділів з'являються гаплоїдні клітини, що дають початок вельми скороченим за розміром макро- і мікрогаметофіту.
Мейоз протікає схоже майже у всіх організмів (рис. 3.4). Два поділу мейозу умовно називають мейоз I і мейоз П. В кожному поділі мейозу, як і в мітозі, розрізняють профазу, метафазу, анафазу і телофазу. Реплікація хромосом здійснюється в періоді S - интерфазе, що передує мейозу I. На цій стадії діляться клітини ще не детерміновані до мейозу і можуть знову ділиться мітотично, у них простою зміною живильного середовища можна індукувати або митотичні, або мейоті- етичні ділення. Критичною стадією, на якій клітина незворотно втягується в мейоз, є профаза мейозу I. Дріжджові клітини на цій стадії вже не можуть повернутися до мітотичним контрольними позначками і гинуть, якщо перервати нормальний перебіг профази мейозу.
Профаза I - складно організована стадія. Її прийнято поділяти на п'ять етапів: лептотене, зіготене, пахітене, ді плоті ну і діакінеза (рис. 3.4).
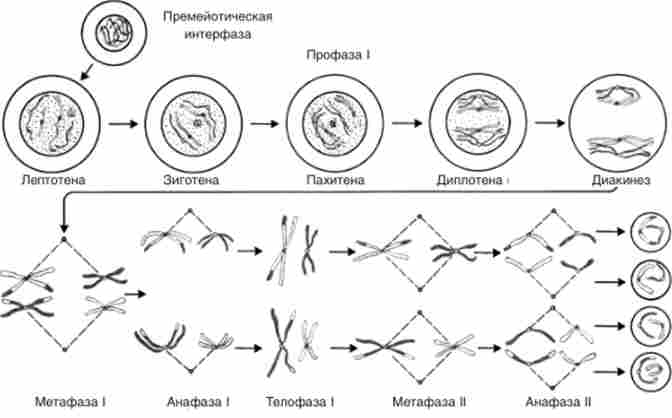
Мал. 3.4. Схематичне зображення послідовних стадій мейозу
Послідовні стадії мейозу були вперше детально досліджені Уініуортером (1900) при вивченні яєчників кролика, оскільки у цієї тварини все стадії мейозу при утворенні ооцитів дуже розтягнуті.
Лептотена (Стадія тонких ниток) нагадує ранню профазу мітозу. На цій стадії з'являються тонкі перекручені нитки хромосом. Іноді виділяють і стадію пролептотени - період, коли хромосоми вже подвоєні, але сестринські хроматиди важкорозрізнювані.
Зіготепа знаменується кон'югацією окремих ділянок гомологічних хромосом, яка завершується по всій їх довжині до кінця зиготи. На стадії зиготи у багатьох тварин утворюється так звана фігура букета, а у рослин відбувається сінезіс, т. Е. Стиск хромосом в клубок. В обох випадках хромосоми орієнтуються теломерной кінцями до одного з полюсів ядра.
Для цієї стадії характерно поява сінаптонемного комплексу (СК), що входить до складу бивалента, - пари кон'югується хромосом. В електронному мікроскопі СК має вигляд трьох або двох електронно-щільних тяжів, розташованих між кон'югується хромосомами. СК - неодмінний атрибут кон'югації хромосом в мейозі.
викрадена (Стадія товстих ниток) характеризується гаплоидним числом бівалентов, т. Е. Фігур, утворених кон'югується хромосомами, кожна з яких складається з двох хроматид. Число бівалентов, таким чином, так само гаплоидному числу (л) хромосом. На цій стадії добре помітний хромомерное малюнок хромосом. У пахітене завершується формування сінаптонемного комплексу. У пахітене і зиготене відбувається невеликий додатковий синтез ДНК - близько 0,3 і 0,1% відповідно. Реплікація ДНК пов'язана з відтворенням хромосом, відбувається в періоді S клітинного циклу (в інтерфазі).
В ді тіло ж не найбільш чітко видна структура бівалентов і складові кожен з них чотири хроматиди. На цій стадії починається відштовхування гомологів і стають помітними фігури, що нагадують грецьку букву X, так звані Хіазм, які свідчать про обмін в біваленте гомологічними ділянками хромосом. У діплотене помітна велика спирализация хромосом, ніж на стадії пахітени.
У тварин в ядрах дозрівають ооцитів на цій стадії спостерігаються характерні утворення - хромосоми відходять від них бічними петлями, на яких в свою чергу розташовані нитки різної довжини. Це хромосоми типу лампових щіток. Помітні обернено пропорційні відносини між величиною хромомер і петель. Це узгоджується з уявленням про те, що петлі «лампових щіток» - результат деспіра- лизации певних хромомер.
У діплотене утворюються численні ядерця (до 1 000 на ядро). Дсспіралізація хромосом, мабуть, відповідає прояву їх метаболічної активності, оскільки на даній стадії дозрівання ооцита виробляються продукти активності хромосом «про запас». Це необхідно у зв'язку з тим, що подальший розвиток плідного яйця відбувається за рахунок цитоплазми матері. Гени, внесені сперматозоїдом при заплідненні, часто не функціонують до стадії гаструли.
В діакінезе спирализация посилюється, зменшується число хіазм, біваленти розташовуються по периферії ядра.
метафаза I. Руйнується ядерна мембрана, і профази змінюється Метафаза. Зникають ядерця. Біваленти розташовуються в екваторіальній площині клітини, утворюючи метафазну пластинку. Хромосоми при цьому сильно спіралізують - потовщені і вкорочені. Спирализация хромосом триває аж до анафази I, коли вони максимально спіралізують.
В анафазе I хромосоми розходяться до протилежних полюсів. Істотна відмінність анафази I мейозу від анафази мітозу полягає в тому, що розходяться хромосоми, що складаються з двох хроматид, прикріплених до одного центромере. Батьківська і материнська центромери кожного бивалента розходяться до протилежних полюсів. Центромери різних бівалентов рухаються незалежно один від одного. Відбувається редукція центромер.
телофаза характеризується утворенням ядерної мембрани і відновленням структури ядра.
після нетривалої інтерфази, або інтеркінез, спостерігається другий розподіл мейозу. Від звичайної інтерфази інтеркінез відрізняється тим, що в ньому хромосома подвоюються.
В профазе II хромосоми стають добре помітні. При цьому вони часто виявляються у вигляді фігури хреста, тому що сестринські хроматиди, відштовхуючись один від одного, утримуються не поділили ще центромерой.
метафаза II здійснюється по мітотичного типу. Хромосоми в мейозі відрізняються від хромосом в мітозі на даній стадії: у останніх більш чітко виражена подвійна структура і більше ступінь спирализации. У мітозі дві дочірні хроматиди асоційовані тісніше.
У анафазе П відбувається розбіжність подвоєних центромер, в результаті чого дочірні хроматиди розходяться до різних полюсів.
У телофазе П утворюються чотири гаплоїдних ядра.
При розгляді двох типів поділу клітини не випадково акцентувалася увага на поведінку структур ядра, перш за все хромосом. В даний час тільки для хромосом, кожна з яких в гаплоидном наборі унікальна, відомий механізм точного розподілу в мітозі і мейозі. Для інших клітинних органел, таких як мітохондрії і пластиди, представлених в клітці в безлічі, точний механізм розподілу не вивчений.