| Головна |
| «« | ЗМІСТ | »» |
|---|
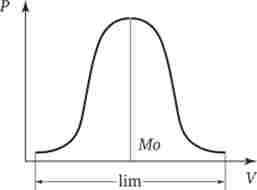
Мал. 7.1. Крива розподілу модифікацій ознаки в варіаційному ряду.
Завдяки впливу факторів середовища на формування фенотипу навіть генетично ідентичні організми в різних умовах розвитку і існування в більшій чи меншій мірі різняться за своїми ознаками. Фенотипічні зміни, що виникають на основі одного і того самого генотипу в різних умовах його реалізації, називають модифікаціями. Прикладом модифікацій можуть служити зміни вмісту жиру в молоці тварин або маси тіла в залежності від їх харчування, зміни кількості еритроцитів в крові, в залежності від парціального тиску кисню в повітрі, зміни темпу росту рослин при різній освітленості і зміст мінеральних речовин в грунті.
V - варіанти ознаки, Р - частота народження варіантів ознаки, Мо - мода, або найбільш часто зустрічається значення ознаки, Ігл - межі модіфікаціон- ної мінливості ознаки (норма реакції)
Модифікації окремого ознаки або властивості, що формується даними генотипом, утворюють безперервний ряд. Частота народження кожного варіанту в такому вариационном ряду різна. Найчастіше виявляються середні значення ознаки. Чим далі ознака відстоїть від середнього значення, тим рідше він спостерігається (рис. 7.1).
У генотипі організму запрограмовано не конкретне значення окремих його характеристик, а лише можливість їх формування в певних межах, які називаються нормою реакції. Таким чином, норма реакції являє собою межі модіфікаці- ційної мінливості ознаки, допустимої при даному генотипі. Деякі ознаки характеризуються широкою нормою реакції. Як правило, це кількісні ознаки, контрольовані полігенами (маса тіла, жирність молока, пігментація шкіри), інші властивості характеризуються вузькою нормою реакції і слабо або майже не модифікуються в різних умовах (колір очей, група крові).
Фенотипічніпрояв інформації, що містяться в генотипі, характеризується показниками пенетрантності експресивності. Пенетрантност' відображає частоту фенотипічного прояви наявної в генотипі інформації. Вона відповідає відсотку особин, у яких домінантний аллель гена проявився в ознака, по відношенню до всіх носіїв цього алеля. Неповна пенетран- тність домінантного алеля гена може бути обумовлена системою генотипу, в якій функціонує даний аллель і яка є своєрідним середовищем для нього. Взаємодія неалельних генів в процесі формування ознаки може привести при певному поєднанні їх алелей до невияв домінантного алеля одного з них.
У розглянутих вище прикладах (див. Розд. 3.6.5.2) наявність в генотипі одного з генів в рецессивном гомозиготномустані не давало можливості проявитися домінантним алелем іншого гена (альбінізм, «бомбейський феномен»). Відомі також випадки, коли фенотипичному прояву певного алеля перешкоджають чинники навколишнього організм середовища. Наприклад, у китайській примули розвиток або відсутність червоного забарвлення квіток залежить від температури і вологості повітря: при t = 5 ^ 20 ° С - червоні квіти, при t = 30-Г35 ° С і підвищеній вологості - білі. У кроликів гімалайської забарвлення темна пігментація вовни, розвивається в звичайних умовах лише на окремих ділянках тіла, при вирощуванні їх при низькій температурі може бути отримана на всьому тілі.
експресивність також є показником, що характеризує фенотипічніпрояв спадкової інформації. Вона характеризує ступінь вираженості ознаки і, з одного боку, залежить від дози відповідного алеля гена при моно- генному успадкування або від сумарної дози домінантних алелів генів при Полигенное успадкування, а з іншого - від факторів середовища. Прикладом служить інтенсивність червоного забарвлення квіток нічної красуні, спадна в ряду генотипів АА, Аа, аа, або інтенсивність пігментації шкіри у людини, що збільшується при зростанні числа домінантних алелей в системі полигенов від 0 до 8 (див. Рис. 3.80). Вплив факторів середовища на експресивність ознаки демонструється посиленням ступеня пігментації шкіри у людини при ультрафіолетовому опроміненні, коли з'являється загар, або збільшенням густоти вовни у деяких тварин в залежності від зміни температурного режиму в різні сезони року.