| Головна |
| «« | ЗМІСТ | »» |
|---|
У першій половині XX в. було встановлено, що порушення від закінчень рухового нейрона передається на м'язове волокно, спонукаючи його до скорочення через спеціальні контакти - нервово-м'язові синапси (рис. 8.14).
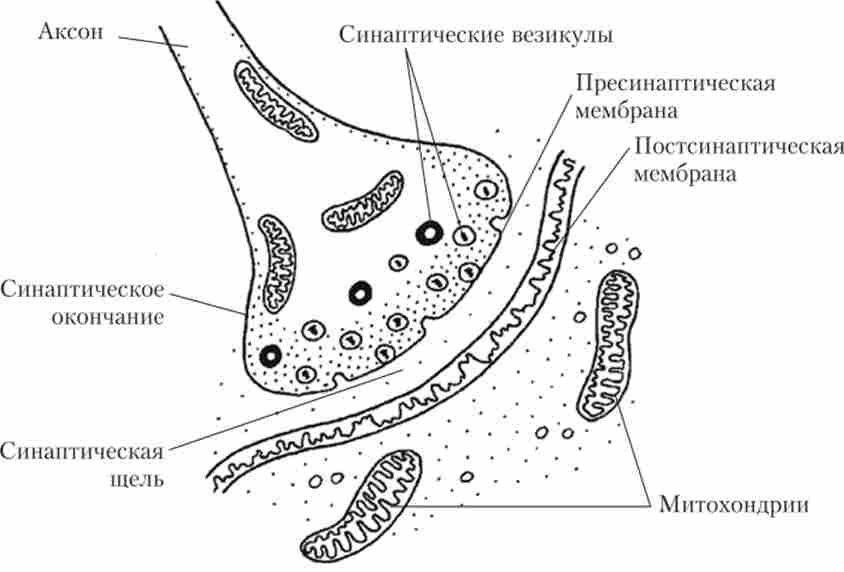
Мал. 8.14. Схема будови хімічного синапсу
Аксон рухового нейрона, підходячи до поперечно-смугастої м'язі, розгалужується і утворює на її поверхні потовщення - синаптичні закінчення. Причому на одному з м'язових волокон закінчується тільки одне розгалуження нервового волокна. Це цілком зрозуміло, тому що величина сигналу, що приходить з одиночного волокна, цілком достатня для того, щоб порушити м'язову клітку і викликати її швидке скорочення. Однак скорочення однієї м'язової клітини не призведе до руху, так як м'язи зазвичай складаються з дуже великого числа клітин. Тому аксон мотонейрона галузиться в м'язі, утворюючи закінчення (синапси) на декількох м'язових волокнах. Виходить, що один руховий нейрон (мотонейрон) управляє багатьма м'язовими волокнами. Мотонейрони і ті м'язові волокна, якими цей мотонейрон управляє, разом називаються руховою одиницею (Рис. 8.15).
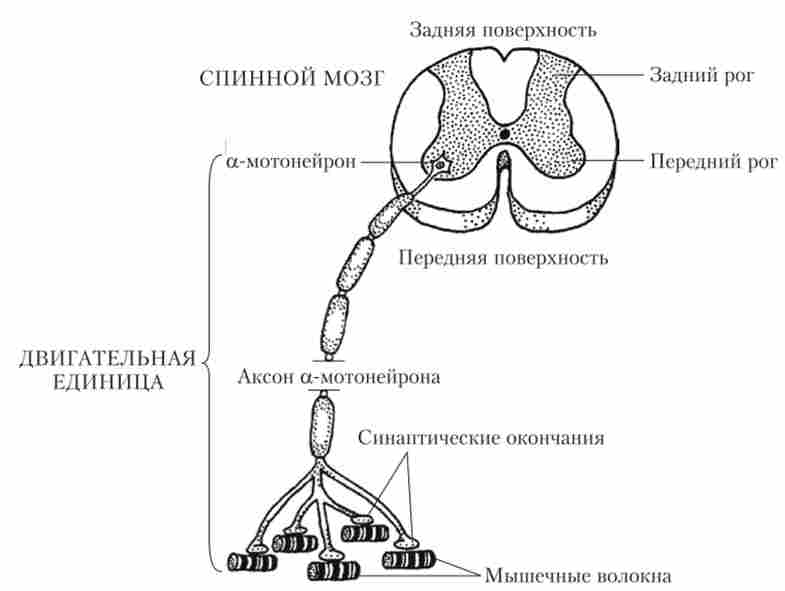
Мал. 8.15. Схема рухової одиниці
В очних м'язах, що виконують дуже точні рухи, один мотонейрон управляє всього двома-п'яти м'язовими волокнами, т. Е. Рухова одиниця дуже маленька. У м'язах пальців руки в рухової одиниці містяться 10-20 м'язових волокон. У литкового м'яза, яка не повинна вчиняти дуже точних рухів, рухова одиниця включає до 1000 волокон.
Повернемося до пристрою нервово-м'язового синапсу. Синаптичні закінчення дуже малі: їх діаметр 1-2 мкм. У пресинаптичному закінчення знаходяться мікроскопічні бульбашки, стінки яких утворені мембраною. Усередині цих бульбашок, або, як їх частіше називають, синаптичних везикул, знаходяться молекули медіатора - хімічного передавача збудження від нервового закінчення на мембрану м'язової клітини. У 1936 р було показано, що в нервово-м'язовому синапсі медіатором є АХ, хоча в НС в контактах між нейронами в різних відділах мозку виявлено велику кількість медіаторів. В одному бульбашці знаходиться кілька тисяч молекул АХ. Порцію молекул медіатора, що міститься в одній везикули, називають квантом медіатора, причому в одній і тій же різновиди синапсу кванти практично однакові. Наприклад, у теплокровних тварин, а значить, і у людини, квант АХ становить 2 - 104-4 - 104 молекул.
Який же механізм викиду АХ в синаптичну щілину? По-перше, необхідно деполярізовать мембрану пресинаптичного закінчення на 30-40 мВ, що і відбувається, коли по нервовому волокну до оконча-
нию підходить ПД. По-друге, в цей момент в мембрані цього закінчення відкриваються Са2+-канали і Са2+ надходить в закінчення з міжклітинної середовища (рис. 8.16).
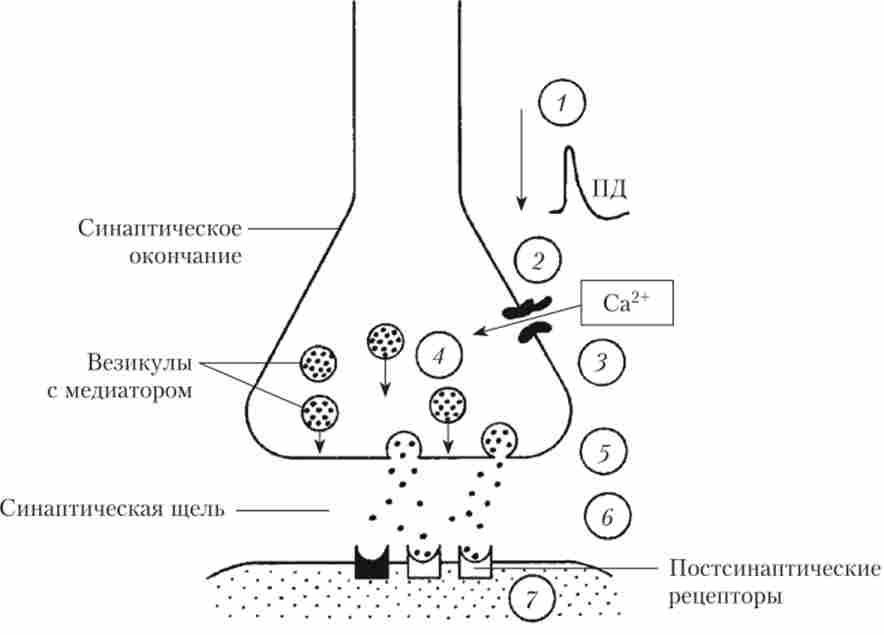
Мал. 8.16. Етапи синаптичної передачі:
Кальцій необхідний для взаємодії особливих білків, які роз- положення в мембранах везикул і пресинаптичного закінчення і забезпечують зближення мембрани везикул з пресинаптичної мембраною. Тепер Везикула може на мить підійти до мембрани закінчення, вбудуватися в неї і викинути квант медіатора в синаптичну щілину. Далі необхідно дуже швидко видалити з закінчення надлишок кальцію проти градієнта концентрації. Це завдання виконують спеціальні насоси, що представляють собою білкові молекули. Увійшовши в синаптичну щілину, молекули АХ зв'язуються з рецепторами, розташованими на постсинаптичні мембрані, і в цей момент проникність ділянки мембрани, що лежить під синаптичним закінченням, різко збільшується; в постсинаптичну клітку з міжклітинної середовища спрямовуються Na+ і Са2+. Зусиллями багатьох дослідників було показано, що рецептори до АХ, розташовані на клітинах скелетних м'язів, є частиною іонного каналу (рис. 8.17).
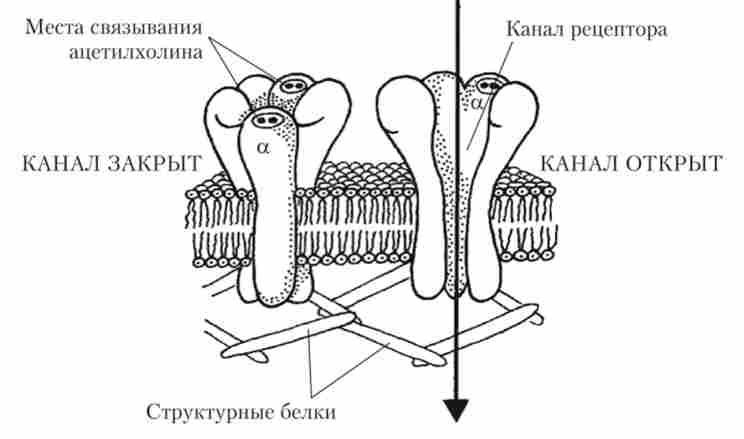
Мал. 8.17. Тривимірна модель нікотинових ацетилхолінових рецепторів:
рецептор складається з п'яти білкових субодиниць, з яких дві ідентичні (а) мають ділянки зв'язування з АХ. Якщо обидві ділянки зв'язування зайняті АХ, відкривається іонний канал
П'ять білкових субодиниць, розташованих в постсинаптичні мембрані під нервовим закінченням, утворюють канал. Дві молекули АХ зв'язуються зі спеціальними ділянками на двох однакових субодиниць, що позначаються а. В цю мить просторова структура всього каналу змінюється, і по ньому всередину постсинаптичної клітини надходять Na+ і Са2+. На відміну від каналів для Na+, розташованих в мембрані аксона, що регулюються змінами потенціалу на мембрані (т. е. потенціалзавісімих), описуваний канал назвали хемовозбудімим катіонних, оскільки він пропускає виключно катіони, а його «ворота» відкриваються хімічною речовиною (в даному випадку - АХ). В результаті надходження в постсинаптичну клітку позитивно заряджених іонів мембранний потенціал зсувається до нуля, т. Е. Спостерігається деполяризація. Якщо ця деполяризація досягне критичної величини, при якій починають відкриватися потенціалзалежні Ка+-канали, то постсинаптична клітина збудиться - в ній виникне ПД. У разі нервово-м'язового синапсу так і буває, а ось нервова клітина рідко деполяризуется до критичного рівня під дією одиночного сигналу, що прийшов через синаптичних контакт.
Молекула АХ взаємодіє з рецептором менше 1 мс. Якщо взаємодія триває довше - рецептор робиться рефрактерним і надовго виходить з ладу. Отже, необхідно швидко руйнувати АХ в синаптичної щілини. Для цього в синаптичної щілини присутній фермент ацетилхолінестерази. У кожній молекулі цього ферменту є до 50 активних центрів, і одна молекула здатна зруйнувати до 17 млн молекул АХ. Утворені в результаті цієї реакції холін і ацетат
захоплюються спеціальною системою активного транспорту і поглинаються нервовим закінченням для вторинного використання при синтезі нових порцій АХ.
Коротко перерахуємо явища, що відбуваються при роботі нервово-м'язового синапсу.
Отже, очевидно, що передача збудження з нерва на м'яз - процес складний, що включає в себе кілька етапів, і в той же час дуже важливий для будь-якого організму: надійність і швидкість синаптичної передачі багато в чому визначають швидкість рухів, від якої часто залежить саме життя тваринного .
Тому немає нічого дивного, що в процесі еволюції з'явився цілий ряд отрут - блокаторів нервово-м'язової передачі на тому чи іншому етапі цього процесу. Адже хижак, який своїм укусом паралізує жертву, голодним не залишиться.
Вище вже говорилося, що лідокаїн і його аналоги місцевої дії блокують Иа+-канали в мембрані нервових волокон, в тому числі і рухових волокон, придатних до скелетних м'язів. В результаті амплітуда ПД знижується, закінчення нервового волокна деполяризуется слабо і АХ з везикул не виходитиме в синаптичну щілину.
Ряд отрут блокує викид АХ з везикул в синаптичну щілину, в результаті чого м'язи перестають скорочуватися, розвиваються паралічі, а потім зупиняється дихання. Прикладом таких отрут можуть служити p-бунгаротоксін (отрута змії смугастого крайта), нотексін (отрута тигровій змії), тайпоксін (отрута змії тайпан) і ін. Дуже страшний токсин виділяють бактерії ботулізму (Clostridium bolulinutri). Їх отрута - один з найсильніших: всього 0,5 мкгтоксіна вбиває людину. Ці бактерії живуть в грунті. В організмі людини токсин ботулізму потрапляє з неправильно консервованими продуктами, в які потрапили суперечки цих бактерій. Потрапили в організм бактерії виділяють токсин, який перешкоджає контакту везикули (з АХ) і мембрани пресинаптичного закінчення - АХ не викидається в синаптичну щілину. Приблизно через дві доби після отруєння в очах починається двоїння, порушується мова, розвиваються паралічі, зупиняється дихання.
Зовсім інший механізм дії у отрут змій, що відносяться до бунгарусам: а-бунгаротоксін «сідає» на рецептори а-субодиниць катіонного каналу, т. Е. На ті місця, з якими повинен був взаємодіяти ЛХ. Причому его взаємодія отрути і рецепторів необоротно, і якщо тварина виживає, то рецептори до АХ в м'язах доводиться «виготовляти» заново.
Найбільш відомим з отрут - блокаторів нервово-м'язової передачі - є б / -тубокурарін (кураре). Ця отрута, що виділяється з насіння рослин сімейства луносемян- іікових, так само блокує постсинаптичні рецептори до АХ, витісняючи медіатор і тимчасово (оборотно) припиняючи роботу синапсу.
Блокада ацетил холінестерази (АХЕ) в синаптичної щілини призведе до накопичення АХ, посиленому впливу на рецептори і, як наслідок, судом і загибелі організму. Найвідомішим з природних блокаторів АХЕ є фізостигмін, що виділяється з західноафриканської рослини калабарских боб (сімейство бобові).
Штучно були створені десятки нервово-паралітичний отрут, які блокують АХЕ і порушують в першу чергу роботу мозку, а потім вже нервово-м'язову передачу. До цих отрут відносяться зарин, замало, табун, VX.
Відомі також захворювання, що вражають нервово-м'язову передачу. Одне з них - міастенія. При цьому захворюванні, що носить генетичний характер, відбувається руйнування рецепторів до АХ або інших компонентів синапсу аутоантителами (т. Е. Антитілами до власних білків організму). Для того щоб посилити роботу поки що непошкоджених синапсів, хворим вводять малі дози фізостигміну, блокуючи АХЕ і продовжуючи існування АХ в щілини для того, щоб молекули медіатора встигали зв'язатися з рештою рецепторами і провести сигнал від мозку до м'язів для забезпечення їх нормального скорочення.