| Головна |
| «« | ЗМІСТ | »» |
|---|
В основі сучасного уявлення про структуру та функції ЦНС лежить нейронна теорія. Нервова система побудована з двох типів клітин: нервових і гліальних, причому число останніх в 8 ... 9 разів перевищує число нервових. Однак саме нейрони забезпечують все різноманіття процесів, пов'язаних з передачею та обробкою інформації.
Таким чином, структурно-функціональна одиниця ЦНС - нервова клітина, або нейрон. Окремі нейрони, на відміну від інших клітин організму, що діють ізольовано, «працюють» як єдине ціле. Їх функції полягають у передачі інформації (у формі сигналів) від однієї ділянки нервової системи до іншого, в обміні інформацією між нервовою системою і різними ділянками тіла. При цьому передавальні і приймаючі нейрони об'єднані в нервові мережі і ланцюги.
У нервових клітинах відбуваються складні процеси обробки інформації. З їх допомогою на зовнішні і внутрішні подразнення формуються відповідні реакції організму - рефлекси.
Нейрони мають ряд ознак, загальних для всіх клітин тіла, незалежно від місцезнаходження і функцій. Будь-нейрон має плазматичну мембрану, яка визначає межі індивідуальної клітини. Нейрон взаємодіє з іншими нейронами або вловлює зміни в локальному середовищі за допомогою мембрани і укладених в ній молекулярних механізмів. Варто відзначити, що мембрана нейрона значно міцніше, ніж інших клітин.
Все, що укладено всередині плазматичноїмембрани (крім ядра), називається цитоплазмою. Тут містяться цитоплазматичні органели, необхідні для існування нейрона і виконання ним своєї роботи. Мітохондрії забезпечують клітину енергією, використовуючи цукор і кисень для синтезу спеціальних високоенергетичних молекул, які витрачаються кліткою у міру потреби. Микротрубочки-тонкі опорні структури - забезпечують підтримку певної форми нейрона. Мережа внутрішніх мембранних канальців, за допомогою яких клітина розподіляє хімічні речовини, необхідні для її функціонування, називається ендоплазматичним ретикулумом.
Розрізняють два види ЕПР: шорсткий і гладкий. мембрани шорсткого (Гранулярного) усіяні рибосомами, необхідними клітинам для синтезу секретується ними білкових речовин. Велика кількість елементів шорсткого ретикулума в нейронах характеризує їх як клітини з дуже інтенсивною діяльністю. Інший вид плазматичного ретикулума - гладкий, званий також апаратом Гольджі, упаковує речовини, синтезовані кліткою, в спеціальні «мішечки», побудовані з мембран гладкого ретикулума. Завдання цієї органели нейрона полягає в перенесенні секретів до поверхні клітини.
Мал. 13.1. Схематичне зображення нейрона (пропорції змінені):

/ - дендрити; 2-тіло клітини; 3 - аксонний горбок; 4 аксон; 5 - коллатераль аксона; 6 - місліновая оболонка; 7- шваннов- ська оболонка; 8- ядро шванівської клітини; 9- перехоплення ран- вье; 10 кінцеві розгалуження аксона, позбавлені мієлінової оболонки
У центрі цитоплазми знаходиться ядро, в якому, як і у всіх клітин з ядрами, міститься генетична інформація, закодована в хімічній структурі генів. Відповідно до цієї інформацією повністю сформована клітина синтезує специфічні речовини, які визначають її форму, хімізм і функцію. Однак на відміну від більшості інших клітин тіла зрілі нейрони не можуть ділитися. Тому генетично обумовлені хімічні елементи будь-якого нейрона повинні забезпечувати збереження і зміна функцій протягом усього його життя. У великих нейронах ядро займає чверть або третина їх тіла. Вхідні в його склад ядерця беруть участь у постачанні клітини рибонуклеїнова кислота і білками (в мотонейронах, наприклад, при рухової активності тварини ядерця значно збільшуються в розмірах).
Разом з тим нейрони на відміну від інших клітин організму крім тіла (соми) мають відростки (рис. 13.1). Численні чутливі короткі древовидно-розгалужені відростки - дендрити (гр. Дерево) служать своєрідними входами, через які сигнали надходять в нервову клітину. Вони мають шорстку поверхню, обумовлену невеликими потовщеннями - шипиками, немов намистинки, нанизаними на дендрит. Завдяки цьому збільшується поверхня нейрона і максимально підвищується збір інформації.
Виходом нейрона є відходить від гена довгий гладкий відросток - а до с о н (від гр. Axis - вісь), який передає нервові імпульси іншій нервовій клітині або робочого органу. Аксони багатьох нейронів покриті мієлінової оболонкою, утвореної шванновскими клітинами, багаторазово (10 ... 65 шарів) «намотаних» подібно ізоляційної стрічки навколо стовбура аксона. Однак муфти шванновских клітин, надіті на аксон, не стикаються один з одним. Між ними залишаються вузькі щілини - перехоплення Ранвье, завдяки яким нервове волокно безпосередньо стикається з позаклітинної рідиною. Тому в нервовій системі ссавців тварин хвиля нервового імпульсу поширюється не плавно, а стрибками (сальтаторно) від одного перехоплення до іншого, що дуже прискорює процес просування імпульсу.
Початкова частина аксона - початковий сегмент в місці виходу з тіла клітини (область «аксонного горбка») позбавлена міеліно- виття оболонки, а його мембрана володіє високою збудливістю. Тому її називають пусковий зоною, так як саме звідси починається збудження нейрона.
Для здійснення внутрішньомозкових зв'язків потрібні дуже довгі відростки; аксони виходять за межі ЦНС і прямують до м'язів, залоз, внутрішніх органів. Зібрані в пучки, вони утворюють нерви. Довжина аксонів, що йдуть на периферію, залежить від розміру тварини: у великої собаки - 0,5 м, у коня - більше 1, у жирафа і слона -3, а у гігантського кальмара - 8 м.
Якщо нейрон утворює вихідні зв'язку з великим числом інших клітин, то його аксон може багаторазово галузитися, щоб сигнали могли дійти до кожної з них: кількість таких розгалужень (терміполей) величезна - від 1000 до 10000 і більше. Крім того, аксон здатний віддавати додаткові гілки - колатералі, по яких збудження йде далеко в сторону від магістрального шляху. Цікавий наступний факт - відростки, роз'єднані з тілом клітини, довго існувати не можуть і гинуть. Тіло клітини, навпаки, регенерує їх. Звичайно, це відноситься тільки до центральної частини відростка. Іноді процеси регенерації відростків йдуть з величезною швидкістю: до 30 мк в 1 хв.
Слід зазначити, що саме через наявність відростків нейрони як клітини були відкриті пізніше інших клітин, так як в поле зору мікроскопа нейрон з усіма своїми відростками поміститися не міг. Тому спочатку самим клітинам не надавали належного значення, розглядаючи їх як потовщення серед безлічі відростків.
Форма нейрона, розмір, розташування відростків різноманітні і залежать від функціонального призначення. Кожен окремий нейрон унікальний і нерівноцінний собі подібним на відміну від інших клітин організму. Розміри нейронів дуже варіабельні: найбільші в десятки і сотні разів більше самих дрібних. Наприклад, діаметр зернистих клітин мозочка складає 7 мк, а моторних нейронів спинного мозку - 70 мк.
Щільність розташувань нейронів в деяких відділах ЦНС дуже велика. Так, в корі великих півкуль вона становить 40 000 клітин в 1 мм3. Вважається, що мозок людини і високоорганізованих тварин містить приблизно десятки мільярдів нейронів.
Класифікація нейронів. Сучасна нейробиология пропонує дві класифікації нейронів.
Згідно з першою все нервові клітини діляться на три типи: аферентні (доцентрові, сенсорні, чутливі), еферентні (відцентрові) і проміжні (вставні, контактні, асоціативні, інтернейрони).
Аферентні нейрони передають імпульси (інформацію) від рецепторів в ЦНС. Тіла їх розташовані поза ЦНС - в спинномозкових або черепно-мозкових гангліях (поруч з головним і спинним мозком). Аферентні нейрон має псевдоуні- полярну форму, т. Е. Обидва його відростка виходять з одного полюса клітини. Один з відростків направляється на периферію, де закінчується рецептором (аксоноподобний дендрит), а інший - в ЦНС (істинний аксон). До аферентні нейронам також відносяться нервові клітини, аксони яких складають висхідні шляхи головного і спинного мозку.
Еферентні нейрони працюють в відцентровому режимі, т. Е. Вони пов'язані з передачею низхідних імпульсів від вищерозташованих поверхів нервової системи до ніжерасполо- женним. Наприклад, від кори до спинного мозку або від спинного мозку до робочих органів. Для еферентних нейронів характерні розгалужена мережа дендритів і один довгий аксон. Необхідно відзначити, що число еферентних нейронів в 4 ... 5 разів менше аферентних.
Проміжні нейрони, як правило, більш дрібні клітини, що здійснюють зв'язок між різними нейронами (зокрема, аферентні і еферентних). Вони передають нервові імпульси в різних напрямках (горизонтальному, вертикальному) по ЦНС. Завдяки численним розгалуженням аксона проміжні нейрони можуть одночасно порушувати велика кількість інших нейронів. В ЦНС переважають проміжні нейрони (рис. 13.2).
За другою класифікацією всі нейрони діляться на збуджуючі і гальмівні.
Нейроглія. Безперечно, що нервові клітини - головні трудівники мозку, але їх в мозку значно менше, ніж інших клітин. Виявляється, для здійснення процесів нормального харчування та обміну речовин нейронів необхідні численні клітини нейроглії (гліальні клітини). У мозковій тканині вони заповнюють весь простір між нейронами: в корі великих півкуль їх приблизно в 5 разів більше, ніж нейронів.
Розрізняють три типи клітин нейроглії: астроцити (зіркоподібні, з великою кількістю відростків); олігодендро- ціти (округлі або багатокутні з малою кількістю відростків); мікроглія (дрібні клітини різноманітної форми).
Клітини нейроглії щільно оточують всю капілярну мережу в мозковій тканині; вільна від них лише незначна частина поверхні судин (близько 15%). Вирости гліальних клітин можуть
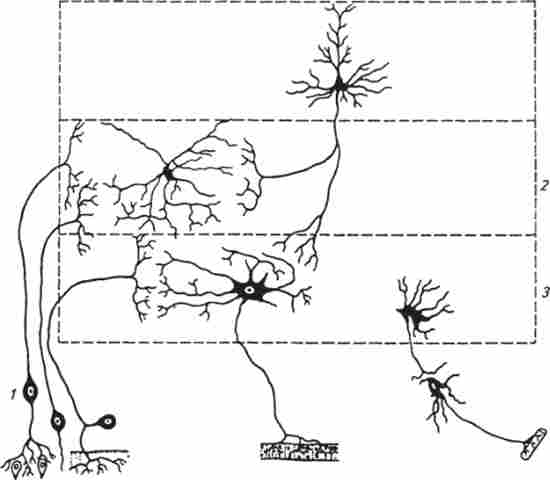
Мал. 13.2. Типи нервових клітин:
/ - нейрони; 2 проміжні нейрони; / -Ефферентние нейрони
розташовуватися з одного боку на нейроні, з іншого - на кровоносних судинах, що вказує на їх участь в передачі поживних речовин і кисню з крові в нервову клітину. Експериментально доведено, що нейроглия активно підтримує функціонування нейрона. Наприклад, при тривалому порушенні високий вміст білка і нуклеїнових кислот в нейроні підтримується за рахунок клітин глії, в яких рівень цих речовин відповідно падає. У процесі ж відновлення після роботи запаси білка і нуклеїнових кислот спочатку наростають в клітинах цієї глії, а лише потім в цитоплазмі нейрона. Цікаво відзначити, що нейрогліальні клітини дуже мобільні і можуть переміщатися в напрямку до більш активних нейронів. Таким чином, в разі необхідності компенсується доставка поживних речовин і кисню до активно «працюють» нейронам.
Клітини нейроглії, імовірно, виконують функцію своєрідної гідродинамічної подушки, що оберігає чутливі і ніжні освіти нейронів від різних фізичних впливів. Останнім часом з'явилися повідомлення про участь цих клітин в умовно-рефлекторної діяльності мозку і механізмах пам'яті.
Таким чином, система нейрон - нейроглії постійно знаходиться в стані гнучкого ритмічно коливається рівноваги. Але в цих процесах, що відбуваються в мозку, безумовно, переважають нейрони, використовуючи нейроглії для підтримки свого функціонування.
Синапси в ЦНС. Діяльність мозку неможлива без взаємодії нейронів один з одним. Труднощі в передачі збудження полягає в тому, що кожна нервова клітина, одягнена в власну оболонку, оточена нейроглії і закутана шарами мієліну, являє собою маленьке самостійне утворення.
Для обміну інформацією необхідні спеціальні пристрої. Ці пристрої - місця контакту двох нейронів - назвав синапсами (від гр. Synapsis - зв'язок, з'єднання, змикання) в кінці XIX в. англійський фізіолог Чарльз Шеррингтон. Правда, в той час ще не було відомо, що нервові клітини обмінюються між собою за допомогою хімічних передавачів. Такі сигнали в подальшому були названі хімічними (вони переважають в мозку ссавців). Крім них в ЦНС, в основному у нижчих тварин, зустрічаються електричні (ефапси) і змішані синапси. Хімічні синапси складаються з пресінапті- чеський і постсинаптичної мембран (рис. 13.3). Пресінапті- чна мембрана містить невеликі пори, здатні пропускати медіатор, який як би упакований в крихітних синаптичних бульбашках діаметром 20 ... 60 нм, локалізованих поблизу мембрани. Виділення медіатора стає можливим завдяки тому, що синаптичні бульбашки «зсуваються» нервовим імпульсом в область так званої активної або оперативної зони. Чим більше нервових імпульсів проходить через синапс, тим більше бульбашок накопичується в цій зоні і прикріплюється до пресинаптичної мембрани. В результаті цього полегшується виділення медіатора при наступних нервових імпульсів.
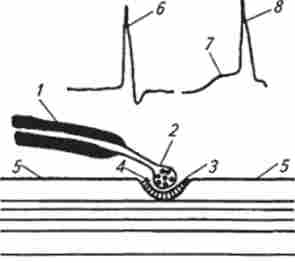
Мал. 13.3. Схема синаптичного зв'язку:
/ -Нервово волокна; 2 нервові волокна з бульбашками медіатора, 3 постсинаптична мембрана; 4-синаптична шсль; 5 - прссінаптічес- кая мембрана м'язового волокна; б-потенціал дії нервового волокна; 7-потенціал кінцевої пластини; 8 - потенціал дії м'язового волокна
У кожному бульбашці пресинаптичної мембрани міститься кілька тисяч молекул медіатора. При цьому деякі пори мембрани прикриті нещільно і бульбашки постійно (по одному в секунду) виливаються в синаптичну щілину. Однак вивільнення такої незначної порції медіатора залишається без відповіді, так як не вистачає критичної маси речовини для здійснення тих процесів, які повинні розвиватися.
Відомі два типи хімічних синапсів, що розрізняються за характером процесів, що розвиваються на постсинаптичні мембрані: збуджуючі і гальмівні.
Розглянемо механізм роботи збудливого синапсу. Під час дії нервового імпульсу на нервове закінчення з бульбашок виділяється порція (квант) медіатора і надходить через пори пресинаптичної мембрани в синаптичну щілину. Передбачається, що тільки завдяки вільній дифузії молекули медіатора можуть перетнути цей простір за лічені 1 ... 2 з-3 (Мілісекунда). Як тільки медіатор вступить в контакт з білками-рецепторами в області постсинаптичної мембрани, остання стає проникною для іонів Na+. Ці іони лавиноподібно спрямовуються в клітину і переносять позитивний заряд на внутрішню поверхню мембрани. Після деполяризації постсинаптичної мембрани виникає збудливий постсинаптичний потенціал (ВПСП). При цьому реєструється невелике коливання мембранного потенціалу з амплітудою до 10 мВ. Надалі він переростає в потенціал дії. Необхідність в медіатора відпадає, і він руйнується специфічним ферментом синапсу.
Існують відмінності в будові і функціонуванні синапсів (рис. 13.4):
синаптична щілину гальмівного синапсу вже і становить 20 нм (у збудливого - 30 нм);
постсинаптична мембрана гальмівного синапсу товща і щільна;
пресинаптична мембрана гальмівного синапсу містить менше бульбашок медіатора;
медіаторами в гальмівних синапсах є гліцин, гамма-аміномасляна кислота (ГАМК), іноді - ацетилхолін.
Для включення в роботу т о р м о з н о го синапсу необхідний також імпульс збудження. При досягненні імпульсом пресинаптичної мембрани в синаптичну щілину викидається порція гальмівного медіатора. Діючи як хімічний подразник на мембранні рецептори, гальмівний медіатор незначно змінює проникність мембрани в основному для іонів К+ і С1 ". Оскільки іонів К+ більше всередині клітини, а іонів С1_ - в міжклітинному просторі, вони рухаються в протилежні напрямки: К '- - назовні, С1 ~ - всередину, що збільшує поляризованность мембрани (мембранний потенціал).
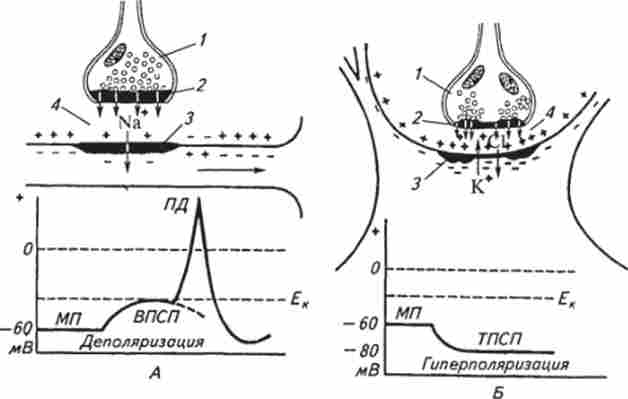
Мал. 13.4. Схема функції збудливого (А) і гальмуючого (Б) синапсів:
/ - бульбашки медіатора; 2 - прссінаптічсская мембрана; 3 постсінаптічсская мембрана; 4 - синаптична щілину; Едо - критичний рівень деполяризації; МП - місцевий потенціал; ВПСП- збудливий і ТПСП - гальмівний постсинаптичні потенціали; ЯЛ - потенціал дії
При цьому реєструється позитивний коливання з амплітудою близько 5 мВ - гальмівний постсинаптичний потенціал (ТПСП). Тривалість ТПСП (як і ВПСП) всього кілька мілісекунд, але протягом цього часу жодне порушення на даному конкретному ділянці постсинаптичної мембрани неможливо. В цьому і полягає механізм дії гальмівного синапсу. Необхідно відзначити, що в деяких нейронах головного мозку тривалість постсинаптичного потенціалу досить значна: ВПСП - до 80 з-3, ТПСП - до 100 з-3.
Беручи до уваги те, що порушення будь-якого нейрона обов'язково супроводжується виникненням біоелектричних потенціалів, виникає питання: чому ж еволюція пішла по більш складного і повільного шляху проведення збудження - через хімічний синапс? Здавалося б, все дуже просто: адже електрику - відмінний подразник, і для передачі збудження з однієї нервової клітини на іншу досить наявності електричного синапсу. Однак мозок не здатний розрізняти потрібне напруга електричного струму, адже нейрон працює за законом «все або нічого», т. Е. Або «мовчить», або дає стандартну відповідь. Там, де збудження обов'язково передається від клітини до клітини, цей принцип можна було б застосувати. Але в мозку найчастіше виникає така ситуація, коли нервова клітина зовсім не відповідає негайним порушенням. В цьому і полягає робота нейронів: постійно аналізувати, чи достатньо ґрунтовна надходить інформація. Саме тому нервові клітини вищих тварин обмінюються між собою інформацією за допомогою хімічних речовин, які легко дозуються.
В мозкової діяльності беруть участь кілька медіаторів: норадреналін, донамін, сербонін викликають збудження; гліцин і гамма-аміномасляна кислота (ГАМК) - гальмування; ацетил холін, універсальний медіатор, - збудження і гальмування. Кожному синапси відповідає тільки один медіатор.
За іншою класифікацією розрізняють такі синапси:
Можливі й інші варіанти. При цьому збудливими, як
правило, бувають аксодендрітічні, а гальмівними - аксоносоматічні і аксо-аксональні синапси.
В ЦНС високоорганізованих тварин частка електричних синапсів (ефапсов) незначна. Передача збудження через ці синапси називається ефаптічну. Пресі- наптічна мембрана в таких синапсах повідомляється з постсинаптичною за допомогою мініатюрних містків, т. Е. Мембрани не розділені щілиною. Потенціал дії, досягнувши пресинаптичної мембрани, перетворюється в градуальний ток, «стікає» на постсинаптичну мембрану, де відроджується як нова хвиля збудження. Таким шляхом збудження проводиться через синапс майже без затримки. Особливість ефапсов полягає в їх здатності передавати збудження в двох протилежних напрямках, що неможливо в хімічних синапсах (рис. 13.5).
У мозковій тканині зустрічаються і змішані сінап- з и. Такий синапс частково може проводити збудження як ефапс або за допомогою медіатора.
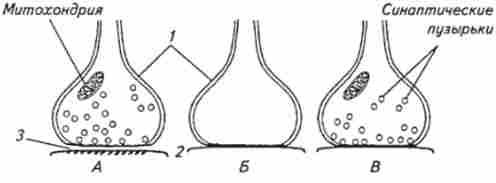
Мал. 13.5. Схема синапсів з хімічними (А), Електричними (Б) і змішаними (В) механізмами передачі:
/ - пресінаптічсская і 2-постсинаптична мембрани; 3 синаптична щілину
Функції нейронів. Нейрон має такими найважливішими функціями: рецепторной - сприйняття різних подразнень; інтегративної - переробка інформації; ефек- битим - передача нервових імпульсів на інші нейрони або орган и-ефектори.
Рецепторная (сприймає) функція нейрона полягає в передачі виникають в рецепторах збуджень в нервову систему через певні ділянки нейрона, що знаходяться в області синаптичних контактів. Відповідь нейронів на роздратування проявляється зміною величини мембранного потенціалу.
Чим більше синапсів на нервовій клітині, тим більше різних подразнень вона сприймає і, отже, тим ширше спектр впливів на діяльність клітини і можливість її участі в різноманітних реакціях організму. Тіло нейрона і значна частина дендритів (50%) буквально всипані синапсами. Особливо густо синапси покривають середню частину і закінчення дендритів, причому багато контактів розташовані на шипики. Так, в мотонейронах спинного мозку і пірамідних клітинах кори поверхню дендритів в 10 ... 20 разів більше поверхні тіла клітини.
Рецепторну функцію виконують аферентні нейрони, що мають велике число різних синапсів (збудливих і гальмівних), здатних до складних процесів переробки надходить в ЦНС інформації.
Інтеграційна функція нейрона - це загальна зміна його мембранного потенціалу в результаті складної взаємодії (інтеграції) місцевих ВПСП і ТПСП всіх численних активованих синапсів на його тілі і дендритах. На мембрані нейрона відбувається процес підсумовування позитивних і негативних коливань потенціалу.
В кінцевому підсумку реакція нервової клітини визначається сумою всіх синаптичних імпульсів. Так, переважання гальмівних синаптичних впливів призводить до гіперполяризації мембрани і гальмування діяльності нейрона. При зсуві же мембранного потенціалу в бік деполяризації збудливість нервової клітини підвищується. Однак електричний розряд нейрона виникає лише тоді, коли зміни мембранного потенціалу досягають порогового значення - критичного рівня деполяризації (близько 10 мВ).
Як вже зазначалося раніше, в нейронах (перш за все великих) збудливість різних ділянок мембрани неоднакова. В області початкового сегмента нервової клітини (аксонного горбка і такий його неміелізірованной частини аксона) знаходиться низькопорогових зона, в якій мембрана нейрона має більш високою збудливістю, ніж на інших ділянках клітини (так, поріг збудливості даної області становить 10 мВ, а області дендрита і інших районів соми - 20 ... 30 мВ). Саме в цій області з моменту досягнення критичного рівня деполяризації починається лавиноподібне рух іонів Na+ в клітку і реєструється потенціал дії (ПД). Тому цю зону прийнято називати пусковий зоною нейрона.
У ПД нервової клітини, так само як і в клітинах інших збудливих тканин (м'язової, залізистої), розрізняють короткочасну високовольтну частину, або пік, і тривалі низькоамплітудні коливання - це слідові потенціали. При цьому необхідно зазначити, що амплітуда піку ПД більшості мотонейронів коливається від 80 до 100 мВ, а його тривалість складає 1,5 с-3.
Еферентна функція нейрона полягає в проведенні збудження від тіла нейрона по аксону до іншої нервовій клітині або робочого органу. У формуванні остаточного еферентної сигналу вирішальну роль відіграють синапси, розташовані ближче до збудливою низькопорогових зоні на тілі клітини (аксоносоматічні синапси). Імпульси, що проходять через ці синапси, як правило, викликають у відповідь розряд нейрона, а імпульси, що діють через віддалені (аксодендрітічні) синапси, можуть привести лише до підпороговим змін його збудливості. Таким чином, аксоносоматічні синапси створюють в основному умови для відповіді нервової клітини на роздратування, але буде відповідь чи ні обумовлюється характером впливів, що надходять через аксодендрітічні синапси від інших нервових центрів.
Таким чином, складаються адекватні реакції, що залежать від багатьох подразнень, що діють на організм в конкретний момент часу. В результаті всього цього здійснюється тонке пристосування організму до мінливих умов зовнішнього і внутрішнього середовища.
Фізіологічні показники функціонального стану нейрона. Функціональний стан нейрона не постійно і може істотно змінюватися. Фізіологічними показниками цього стану є збудливість і лабільність.
Збудливістю нейрона називають його здатність відповідати на синаптичне вплив потенціалом дії. Збудливість визначається співвідношенням двох параметрів - мембранного потенціалу і критичного рівня деполяризації. Треба відзначити, що в нормальних умовах діяльності нейрона критичний рівень деполяризації відносно стабільний (= 10 мВ), тому збудливість нервової клітини в основному залежить від величини мембранного потенціалу. Зазвичай потенціал спокою мембрани нейрона в нормі становить близько 70 мВ, але він може бути зменшений (явище деполяризації) або збільшений (явище гіперполяризації).
Як уже неодноразово зазначалося, для появи ПД при нормальному мембранном потенціал необхідний спочатку оптимальний зсув заряду мембрани в бік деполяризації на 10 мВ
(Амплітуда порогового ВПСП) - ця величина називається порогом збудливості нейрона.
Подивимося, як змінюється збудливість нейрона при зниженні його мембранного потенціалу (це спостерігається в активно працює нервовій клітині або при слабких синаптичних впливах, коли з'являються підпорогової ВПСП). При цьому різниця між рівнем мембранного потенціалу і критичним рівнем деполяризації зменшується. Значить, знижується і амплітуда порогового ВПСП. В результаті для появи відповідного ПД потрібна менша сила подразника. Збудливість нейрона в цьому випадку підвищена.
При зниженні збудливості нервової клітини різниця між рівнем мембранного потенціалу і критичним рівнем деполяризації зростає, і для виникнення ПД потрібно значно більше сильне роздратування (велика амплітуда порогового ВПСП).
Збудливість нейрона у разі порушення змінюється з тієї ж закономірністю, що й в інших клітинах збудливих тканин. У момент появи високовольтної частини - піку (спайка) нейрон не може відповісти на роздратування новим ПД, так як він абсолютно невозбудімості (абсолютна рефрактерність). Ця фаза триває приблизно 0,5 с-3. Потім, в період реполяризації мембрани, збудливість нервової клітини поступово відновлюється до вихідного рівня (відносна рефрактерність). Наступною фазою є екзальтація (підвищення рівня збудливості в порівнянні з вихідним станом), і, нарешті, під час слідової гіперполяризації настає фаза субнормального (незначне зниження рівня збудливості).
Знання закономірностей зміни збудливості нейрона при порушенні має велике значення для розуміння особливостей його діяльності в різних ситуаціях. Так, більшість нейронів проміжного мозку і кори великих півкуль після чергового розряду імпульсів (через відповідної тривалості фаз абсолютної і відносної рефрактерності) неактивно протягом приблизно 100с_3. Отже, найбільш зручний ритм їх спонтанної активності - близько 10 розрядів в 1 с, ніж визначається частота електричних коливань на поверхні мозку в стані спокою.
Лабільність - це швидкість протікання елементарних реакцій, що лежать в основі порушення нервової клітини. Різні за функціями і розмірам нейрони мають різні величинами лабільності: навіть в межах однієї нервової клітини лабільність її різних структур (соми, дендритів, початкового сегмента аксона) різко відрізняється. Величина лабільності нейрона, так само як і збудливості, визначається рівнем його мембранного потенціалу. Лише при певній величині мембранного потенціалу досягається оптимальний рівень збудливості і лабільності нервової клітини, а також найбільш високий рівень її ритмічної активності. Ця обставина, безперечно, є важливою умовою для передачі інформації в ЦНС і здійснення доцільних реакцій. Наприклад, максимальний ритм мотонейронів спинного мозку (до 100 імпульсів в секунду) відзначається при середній, оптимальної величиною мембранного потенціалу - 55 мВ, т. Е. При підвищеній збудливості мотонейронів.
У природних умовах життєдіяльності частота розрядів мотонейрона зазвичай не перевищує 50 ... 60 імп / с, але іноді вона може підвищуватися до 100 ... 300 імп / с. Це буває при потужному впливі вищерозташованих відділів ЦНС і природному зниженні мембранного потенціалу. Однак розряди з такою частотою дуже короткочасні. Вони змінюються більш повільним і стійким ритмом активності.
У дрібних же проміжних нейронах спинного мозку частота розрядів в початкові моменти активації може досягати 600 ... 800 і навіть 1000 імп / с. Це необхідно для різкого і швидкого впливу цих клітин на мотонейрони.
Численні роздратування зовнішнього і внутрішнього середовища, що викликають афферентную імпульсацію, підвищують збудливість і лабільність нейронів, їх здатність до стабільної ритмічної активності, а бездіяльне же стан знижує їх, погіршуючи функціональний стан нервової клітини.
Активна діяльність нейрона викликає не тільки функціональні зміни. Вона веде до значних структурних і біохімічних перебудов його різних елементів (наприклад, до змін в синаптичному апараті нервових клітин). Так, експерименти з тренуваннями тварин (навантаження на різні групи м'язів) показали, що середня величина (діаметр) синапсів на мотонейронах спинного мозку достовірно зростає, приблизно на 35%. Зі збільшенням розмірів синапсів поліпшується проведення нервових імпульсів в ЦНС.
Активну участь коркових нейронів в умовно-рефлекторної діяльності сприяє розвитку шіпікового апарату на дендритах і підсилює міжнейронні взаємозв'язку в корі великих півкуль. Придушення діяльності кори різними фармакологічними препаратами, навпаки, призводить до погіршення рецепторной функції нейронів - порушення синаптичних контактів (зникнення шипиків) і навіть деформації дендритів.
При посиленні афферентной імпульсації, що надходить до нейрона, в синапсах відбувається посилення синтезу медіатора і відповідно збільшення числа синаптичних пухирців, а також їх інтенсивне переміщення в оперативну зону пресінапті- чеський мембрани. Необхідно відзначити, що на тлі посиленої рухової активності можуть збільшуватися навіть розміри ефек фекторних нейронів.
Таким чином, всі структурні, біохімічні та функціональні зміни сприяють здійсненню нервовими клітинами їх складних функцій. Це лежить в основі вдосконалення різних поведінкових реакцій цілісного організму, в тому числі рухової діяльності і психічних процесів.