
| Головна |
| «« | ЗМІСТ | »» |
|---|
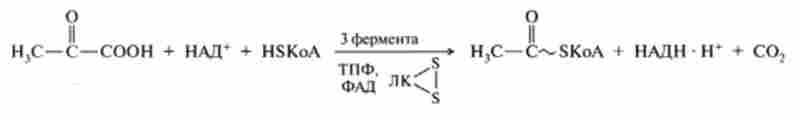
Піруват, що утворився в цитоплазмі клітини, надходить в мітохондрії, де він перетворюється в ацетил-КоА і С02, при дії сложноорганізованного мультиферментного піруватдегідрогеназного комплексу (ПД-комп лскс). До складу ПД-комплексу (табл. 19.1) входять три складних ферменту, ко ферменти яких досить міцно асоційовані з аіоферментамі, і два коферменту - легко диссоциирующие (HS-KoA і НАД *).
Ферменти ПД-комплскса і коферменти, локалізовані в матриксі мітохондрій, об'єднані в Мультиферментний систему, молекулярна маса якої перевищує 6 - 10 * kDa. «Ядро» цього комплексу складає дегідролі- поілтрансацетілаза. до якої приєднані великі молекули двох інших ферментів - піруватдегілрогенази і легідроліпоілдегілрогенази. що дозволяє продукту дії одного ферменту швидко взаємодіяти з іншим.
Таблиця 19.1. Ферменти і коферменти ПД-комплексу
|
фермент Е |
Назва |
кофермент |
функція |
|
Е, |
П іруватлегілрогеназа (ПДГ) |
Гіамін пирофосфат (ТПФ) |
декарбоксилирование пірувату |
|
Дсгід роліпоілтрансацс- тілаза (ДЛТА) |
Ліпосвая кислота (ЛК) |
Перенесення водню і ацетил |
|
|
Ез |
Дс гидрол і поілдс гідрогс- наза (ДДДГ) |
ФАД |
перенесення водню |
|
- |
- |
Коензим A (HS-KoA) |
перенесення ацетил |
|
НАД * |
Перенесення відновлювальних еквівалентів в дихальний ланцюг мітохондрій |
Встановлено, що до складу комплексу входять також ферменти киназа і фосфатаза, виконують регуляторну функцію, механізм якої буде розглянуто нижче.
Кожен фермент ПД-комплексу каталізує певну стадію ОДП.
На першій стадії катализируемой ПДГ (Е,) піруват взаємодіє з коферментом цього ферменту (ТПФ), відбувається активація пірувату, його де- карбоксилирование і освіту оксіетільного похідного ТПФ-Е ,:
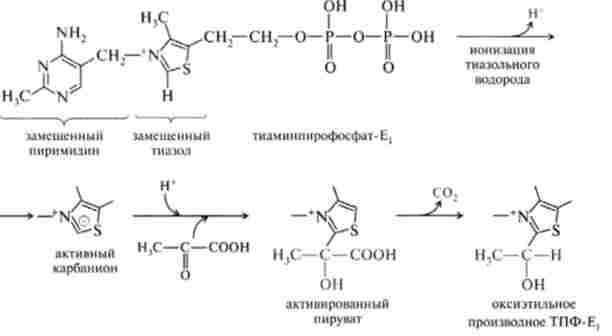
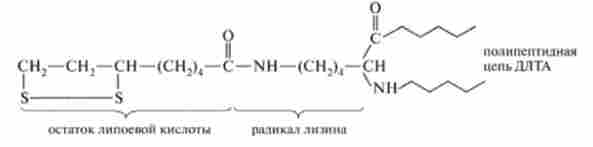
На другій стадії ДЛТА (Е2) Окисляє оксіетільную групу комплексу СН3-СНОМ-ТПФ-Е, і переносить утворюється ацетильную залишок на окислену форму ліпоєвоїкислоти - коферменту ДЛТА (Е2), Який ковалентного пептидного зв'язком приєднаний до залишку лізину пептидного ланцюга ферменту:
s.
Структура окисленої форми ДЛТЛ (| ^ JIK- E2).
s
Нижче наведена реакція, що каталізується Е3:
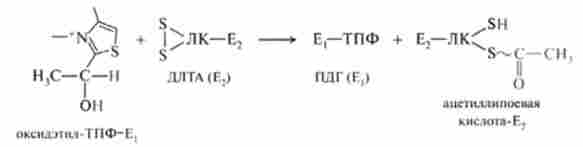
Ацетілліпоевая кислота містить активовану ацетильную групу, втіоефірной зв'язку якої кумулірованнимі енергія окисної реакції дегідрування оксіетільной групи ТПФ.
На третій стадії ДЛТА каталізує перенесення ацетильной групи на HS-KoA. Утворюється один з кінцевих продуктів ОДП-ацетил-КоА і відновлена форма ліпосвой кислоти - коферменту ДЛТА (Е2):
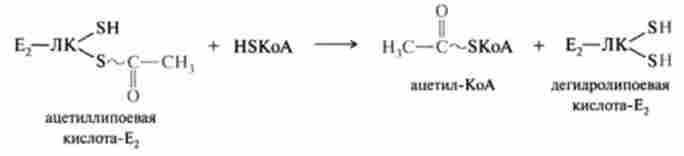
На четвертій і п'ятій стадіях регенеруються окислені форми каталізаторів ПД-комплексу.
На четвертій стадії окислюється дегідроліпоевая кислота, яка входить в з-
sh
ставши відновленого комплексу Е2-Реакція каталізується ДЛДГ (Е3), Що переносить атоми водню від відновлених сульфгідрильних груп дегідроліпоевой кислоти на ФАД-кофермент даного ферменту:
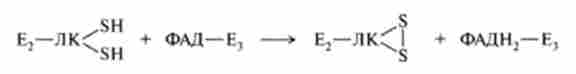
На п'ятій стадії окислена форма ФАДН2-Е3 регенерує за участю НАД+:
Сумарну реакцію ОДП можна записати у вигляді наступного рівняння:
Утворився в процесі ОДП ацетильную залишок у формі ацетил-КоА далі повністю окислюється в ЦТК до С02 і Н20. Слід звернути увагу, що включення ацетильной групи в ЦТК не зажадає витрати АТФ, оскільки в комплексі з коензимом А він знаходиться в активованої формі.
Процес ОДП є енергетично вигідний ще й тому, що в розрахунку на одну молекулу глюкози відбувається відновлення двох молекул НАДН, а отже, в процесі їх окислення дихальної ланцюгом мітохондрій синтезується шість молекул АТФ.
Слід пам'ятати, що окисне декарбоксилювання пірувату є один із загальних шляхів катаболізму, оскільки на рівні пірувату в цей процес втягується ряд метаболітів обміну амінокислот і ліпідів.
Регуляція окисного декарбоксилювання пірувату. Процес, що каталізує ПД-комплексом в тварин тканинах, незворотній, і регуляція його активності становить одну з важливих стадій в регуляції спільних шляхів катаболізму, пов'язуючи між собою такі метаболічні процеси, як гліколіз, глкжонеогенез, синтез і розпад жирних кислот, ЦТК. Як зазначалося вище, до складу ПД-комплексу входять регуляторні ферменти киназа і фосфатаза. Кіназа каталізує фосфорилювання ПДГ (Е,) і переводить цей фермент в неактивний стан, а фосфатаза отшепляет від нього фосфорну кислоту і активує його.
Активність кінази і фосфатази регулюється по аллостерічному механізму. Аллостерічного інгібіторами кінази є піруват, АДФ, HS-KoA, Са2+. Отже, їх висока концентрація підтримує ферменти ПД-комплексу в активній, нефосфорілірованной формі, в клітці створюються умови для освіти з пірувату ацетил-КоА, який може залучатися до ЦТК або використовуватися на синтез жирних кислот.
Аллостерічного активаторами кінази є кінцеві продукти ОДП: ацетил-КоА, НАДН, АТФ. Їх накопичення переводить ПДГ (Е,) в неактивну фосфорильовану форму, припиняється перетворення пірувату в ацетил-КоА, і він може бути використаний, наприклад, для синтезу глюкози.
Отже, ПД-комплекс являє собою складну, саморегулюючу систему, яка відіграє важливу роль як в біологічному контролі дихання і енергетичному забезпеченні організму, так і в регуляції спільних шляхів катаболізму в цілому.