| Головна |
| «« | ЗМІСТ | »» |
|---|
Підшлункова залоза здійснює зовнішню (екзокринну) і внутрішню (ендокринну) секрецію. Клітини, зібрані в складі ацинусів і проток, секретують травні ферменти і виділяють їх в просвіт кишки. Однак в паренхімі органу розташовується особлива тканина, морфологічно оформлена у вигляді острівців і виконує ендокринну функцію. Панкреатичні острівці, що займають від 2 до 10% загальної маси органа, не мають вивідних проток, а виділяють продукти секреції в кров. Острівці, названі по імені описав їх автора острівцями Лангерганса, мають округлу або овальну форму і порівняно невеликі розміри (50 ... 500 мкм). Серед клітин острівців Лангерганса виділяють кілька типів, що розрізняються по величині, формі і забарвленню.
Ацидофільні клітини (А-клітини, а-клітини) розташовуються зазвичай по периферії острівця, мають округлу, рідше полігональну форму. Характерною особливістю А-клітин є наявність в їх цитоплазмі численних секреторних гранул, які представляють собою округлі тільця високою електронної щільності, оточені електронопрозрачним обідком і чітко помітною гладкою мембраною. Секреторні гранули можуть розташовуватися по всій цитоплазмі, але частіше концентруються на полюсі клітини, зверненому до кровоносних судинах. Секретпродуцірующіе структурні компоненти А-клітин представлені незначно шорстким ендоплазматичним ретикулумом: нечисленні канали, вільних рибосом в гіалоплазме трохи, і вони зібрані в невеликі полісомние групи. Комплекс Гольджі виявляється не у всіх А-клітинах, а при наявності цієї структури всередині сплощені бульбашок (в розширених кінцевих відділах) знаходяться локальні згущення матеріалу, ідентичного змісту секреторних гранул. За даними імуногістохімії, А-клітини мають безпосереднє відношення до продукції одного з гормонів підшлункової залози - глюкагону.
Базофільні клітини (В-клітини, (3-клітини) складають клітинну основу, мають полігональну або призматичну форму і займають центральну частину острівця. Цитоплазма В-клітин містить численні секреторні гранули, що відрізняються від відповідних гранул А-клітин своїми розмірами, вони трохи більші за ( діаметр близько 350 нм) і мають меншу електронну щільність. Секреторні гранули розташовуються в секреторних везикулах ексцентрично і оточені більш широким електронопрозрачним обідком, причому мембрана везикули зазвичай має звивистий Перші контури. Морфологічної особливістю секреторних гранул По-клітин у кішок і собак є характерний вид Кристалоїди - прямокутні палички. сек- ретпродуцірующіе структурні компоненти клітин - ендоплазматичнийретикулум і комплекс Гольджі - представлені в В-клітинах досконаліше, ніж в А-клітинах, вони займають велику площу клітини, та й самі клітини значно перевищують А-клітини за своїми розмірами. Дослідження В-клітин у різних видів тварин за допомогою методів мічених атомів і імуногістохімії виявили, що вони обеспечи ают продукцію основного гормону підшлункової залози - інсуліну.
Дефінітивного клітини (Д-клітини, 5-клітини) в острівцях зустрічаються досить рідко, розташовуються поодинці або невеликими групами, мають довгі цитоплазматичні відростки, що надають їм зірчасті форму. Характерною особливістю Д-клітин є наявність в їх цитоплазмі специфічних секреторних гранул, що відрізняються за своєю ультраструктури від А і В-гранул: розміри гранул значно варіюють (200 ... 400 нм), розподілені вони досить рівномірно, іноді концентруються в відростках клітин, звернених до кровоносних судинах. Д-клітини містять нечисленні і слаборозвинені клітинні органели, ендоплазматичнийретикулум представлений невеликою кількістю канальців, комплекс Гольджі розвинений незначно. Функції Д-клітин остаточно не з'ясовані, але дані іммуногістологічнеультраструктурному дослідження дозволяють припускати можливість в них синтезу гастрину, проте не виключається і освіту в них соматостатина, аналог якого продукують нейросекреторні освіти гіпоталамуса.
У ссавців підшлункова залоза виробляє гормон інсулін - поліпептид з молекулярною масою 6000. Для утворення інсуліну особливо необхідний цистеїн, на який в молекулі інсуліну припадає 12 % всього амінокислотного складу. Утворився в В-кліть ках інсулін або відразу надходить в кровотік, або депонується в секреторних гранулах за участю цинку (рис. 12.10). У крові інсулін здатний зв'язуватися з а-, (3- і углобу- лінамі і досягати тканин, які здатні зв'язувати гормон і специфічно реагувати на його присутність. Внаслідок досить швидкого зникнення інсуліну з кровоносноїрусла 20% його виділяється в незміненому вигляді з сечею, решта 80% руйнуються інсуліназой печінки, нирок і м'язів - підшлункова залоза повинна постійно продукувати цей гормон для підтримки певного рівня вуглеводного обміну.
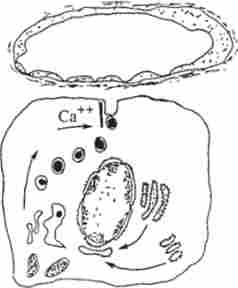
Ріс.12.10. Схема секреторного процесу в інсуліноціте
Інсулін діє на вуглеводний, жировий, білковий і мінеральний обміни, а також на процеси окисного фосфорилювання. В основному його дія проявляється в підвищенні споживання глюкози тканинами, в результаті чого значно знижується вміст цукру в крові. Розвиток гіпбглікеміі при введенні інсуліну пов'язане з тим, що посилюється> тілізація глюкози: дві третини її кількості зв'язується в печінці, а третина в інших органах і тканинах, де глюкоза або згорає з утворенням АТФ, або використовується для синтезу глікогену або ліпідів. Інсулін є єдиним гормоном, який знижує вміст глюкози в крові, тоді як підвищення рівня глюкози може бути викликано декількома гормонами, що утворюються в різних ендокринних органах. При впливі інсуліну посилюється проникнення глюкози в скелетні м'язи через клітинну мембрану, при цьому активується діяльність специфічних переносників. Інсулін здатний брати участь в регуляції обміну білків: стимулює транспорт амінокислот через клітинні мембрани і включення їх в поліпептидні ланцюги, а також підвищує біосинтез білка, посилюючи нуклеїнових обмін (за рахунок інтенсифікації процесів утворення інформаційної РНК і прискорення зчитування інформації з і PH До в момент утворення полісом). Під впливом інсуліну в жировій тканині зростає кількість тригліцеридів, обмін глюкози в ній характеризується переважанням пентозофосфатного шунта - джерела ТРЕБА Н2, який необхідний для синтезу жирних кислот.
Під впливом інсуліну в крові знижується вміст летючих жирних кислот (ЛЖК), які використовуються для утворення жиру в печінці і в інших тканинах, причому в молочній залозі значно збільшується вміст молочного жиру і білків в образующемся молоці.
При нестачі або відсутності інсуліну в організмі виникають серйозні зміни, викликані насамперед тим, що клітини печінки, м'язової тканини перестають отримувати глюкозу з крові за рахунок порушення мембранної транспортної системи. Вміст глюкози в крові значно підвищується, і оскільки гіперглікемія обумовлює підвищення вмісту глюкози в первинній сечі, то нирки не справляються з процесом реабсорбції і частина цукрів виділяється з сечею - розвивається глюкозо Урія ( «цукрове мочеизнурение»). Внаслідок підвищення осмотичного тиску сечі її обсяг збільшується, що призводить до поліурії. В результаті розвивається дегідратація організму, зменшується об'єм циркулюючої крові, знижується артеріальний тиск і порушується мікроциркуляція. Незважаючи на підвищений вміст глюкози в крові тканини за рахунок неможливості її використовувати, відчувають нестачу в джерелах енергії ( «голод серед достатку»).
В ході глюконеогенезу в тканинах починається розпад білків, амінокислоти використовуються для побудови глюкози, рівень якої в крові СШС більш підвищується. Поряд з цим відбувається мобілізація депонированного жиру і, як наслідок, збільшення вмісту в крові жирних кислот, які в печінці перетворюються в кетонові тіла - ацетон, ацстоуксусную і бета-оксимасляную кислоти. Зазвичай ці сполуки можуть використовуватися клітинами, за винятком клітин головного мозку, але при інтенсивному утворенні кетонових тел організм не здатний окислити або метаболизировать їх і кетонові тіла змінюють pH крові, приводячи до ацидозу і вичерпання лужних резервів бікарбонатів плазми. Навіть при посиленому диханні і гіпервентиляції легенів в крові накопичується вуглекислота; якщо pH крові падає нижче 7,0, то розвивається коматозний стан, що веде до смерті. Крім того, ацидоз придушує надходження глюкози в клітини головного мозку, знижується майже в два рази і споживання ними кисню, в результаті порушується синтез АТФ, зменшується вміст кальцію і фосфору, відбувається втрата свідомості, зниження артеріального тиску і ослаблення серцевої діяльності.
Інший гормон підшлункової залози - глюкагон, природний антагоніст інсуліну, має полипептидную структуру, що складається з 29 амінокислотних залишків. Глюкагон (молекулярна маса 3485) бере участь в регуляції вуглеводного обміну за рахунок інтенсифікації розпаду глікогену печінки до глюкози, одночасно пригнічує синтез глікогену і значно підвищує вміст глюкози в крові на противагу інсуліну.
Разом з цим гіперглікемічний ефект посилюється за рахунок глюконеогенезу - перетворення дезамінірованних амінокислот в глюкозу. Вплив глюкагону на ліпідний обмін проявляється в активації ліпаз, що розщеплюють тригліцериди з утворенням вільних жирних кислот. Беручи участь в мінеральному обміні, глюкагон підсилює виведення натрію, калію, кальцію і хлору з сечею і знижує кількість неорганічного фосфату в плазмі крові. На прикладі внутрішньоклітинних ефектів глюкагону вперше була з'ясована роль аденілатциклази і циклічних нуклеотидів (цАМФ) у реалізації дії пептидних гормонів. При активації аденілатциклази і збільшення вмісту внутрішньоклітинної цАМФ відбувається посилення активності фосфорілази печінкових клітин, що призводить до збільшення вмісту глюкозо-1-фосфату і глюкозо-6-фосфату.
Хоча острівці Лангерганса секретують два гормону протилежної дії, наслідки поразки островковой тканини в результаті вірусної інфекції або після видалення підшлункової залози виявляються фатальними за рахунок припинення фізіологічної дії інсуліну, так як глюкагон може крім підшлункової залози синтезуватися в шлунково-кишковому тракті. Крім того, аналогічним глюкагону дією володіють гормон росту, кортизол, адреналін і тироксин.
Порушення транспорту глюкози через клітинні мембрани може відбуватися і при незмінній продукції інсуліну: через порушення рецепторного зв'язування інсуліну клітини гормонзавісі- мих тканин нечутливі до «команді», що виходить із бета-клітин інкреторну відділу підшлункової залози.
Гормони підшлункової залози виділяються в кров безперервно, однак інтенсивність їх надходження може істотно змінюватися відповідно до фізіологічним станом організму. Острівкова тканина знаходиться під контролем вегетативної нервової системи, і парасимпатичні нервові впливу, відтворювані роздратуванням блукаючих нервів, призводять до викиду інсуліну і розвитку гіпоглікемії. Однак денервация залози або її пересадка не призводять до припинення продукції інсуліну, що свідчить про наявність і іншого регуляторного механізму. Основним фактором, що визначає рівень секреції інсуліну, є концентрація глюкози в притікає до підшлунковій залозі крові. При підвищенні вмісту глюкози в крові збільшується виділення інсуліну і знижується секреція глюкагону, причому виділення глюкагону визначається концентрацією цукрів в крові за принципом зворотного зв'язку. Виділення інсуліну в кров може починатися в результаті спрацьовування випереджаючої зв'язку: коли глюкоза ще знаходиться в тонкому кишечнику, місцева ендокринна система шлунково-кишкового тракту до підвищення концентрації цукрів у крові за рахунок впливу секретину і панкреозимина здатна активізувати продукцію інсуліну.
Концентрація глюкози в крові істотно змінюється в залежності від типу їжі. При надходженні в організм вуглеводних кормів вміст інсуліну сприяє відкладенню поживних речовин у вигляді глікогену в печінці і м'язах, жиру - в жирових депо. Гальмування секреції глюкагону і висока концентрація глюкози в крові запобігають глюко- неогенез, і велика частина амінокислот бере участь в білковому синтезі. При переважанні в раціоні білків і дефіциті вуглеводів виникає загроза гіпоглікемії, що особливо небезпечно для тканин мозку, що використовують в якості енергетичного метаболіту глюкозу.
Завдяки секреції глюкагону в цій ситуації підвищується рівень глюкози в крові.
При прийомі їжі, багатої жирами, також необхідна інтенсифікація секреції глюкагону. Активуючи липазу, за рахунок розщеплення тригліцеридів підвищується в крові рівень вільних жирних кислот, які в печінці перетворюються в ацетілКоА і беруть участь в синтезі глюкози. В умовах голодування утворення інсуліну пригнічується, а збільшується вплив глюкагону. Жирова тканина піддається липолизу, вільні жирні кислоти або використовуються безпосередньо в тканинах, або перетворюються в печінці в кетонові тіла, також беруть участь в енергетичному метаболізмі. Розщеплення глікогену до глюкози забезпечує більшу частину потреб тканин мозку, і в подальшому при продовженні голодування для забезпечення поживними речовинами життєво важливих органів при спільній дії глюкагону, адреналіну, АКТГ і кортикостероїдів починається розщеплення білків тканин і утворення глюкози з амінокислот.
Таким чином, інсулін і глюкагон разом здійснюють гормональний контроль обміну речовин. За рахунок зміни співвідношення цих гормонів запобігає марнотратна трата поживних речовин після прийому їжі: зайва глюкоза не виводиться з сечею, а запасається у вигляді глікогену і жирів. Між прийомами їжі співвідношення інсулін - глюкагон встановлюється таким чином, щоб забезпечувалися потреби життєво важливих органів. Інтенсивні м'язові навантаження викликають виділення в кров глюкагону, необхідного для термінового забезпечення м'язової тканини підвищеною кількістю глюкози. Таким чином, при всій протилежності впливу цих гормонів на рівень глюкози синергізм інсуліну і глюкагону забезпечує найбільш повне засвоєння і окислення вуглеводів.
Спільна локалізація інсулін і глюкагон-продукують клітин в межах острівця Лангерганса зумовлює наявність функціонального зв'язку, яка проявляється в тому, що глюкагон здатний стимулювати секрецію інсуліну, а виділяється з Д-клітин соматостатин гальмує секрецію і інсуліну, і глюкагону. Поряд з соматостатином виділення інсуліну гальмують фізіологічно активні речовини симпатичного відділу вегетативної нервової системи і мозкової речовини надниркових залоз - адреналін і норадреналін. У той же час катехоламіни надають стимулюючі впливу на альфа-клітини підшлункової залози, які продукують глюкагон.