| Головна |
| «« | ЗМІСТ | »» |
|---|
Регуляція метаболізму ліпідів становить інтерес насамперед у контексті регуляції енергетичного потоку і шляхи інтеграції його з іншими джерелами енергії в тканинах. Внутрішньоклітинна регуляція процесів окислення і синтезу жирних кислот організована таким чином, що забезпечує першочергове використання в якості енергетичних субстратів вуглеводів і лише в міру їх вичерпання починається окислення жирних кислот (рис. 23.18).
Як видно зі схеми, наведеної на рис. 23.18, якщо в клітини печінки надходить велика кількість глюкози, в результаті піруватдегідрогеназного реакції вона перетворюється в бенкеті ват, карбоксил ювання якого призводить до утворення оксалоацетата. Збільшення концентрації останнього посилює транспорт ацетил-КоА за допомогою нітратного механізму з матриксу мітохондрій в цитоплазму. Цитоплазматичний цитрат активує ацетил-КоА-
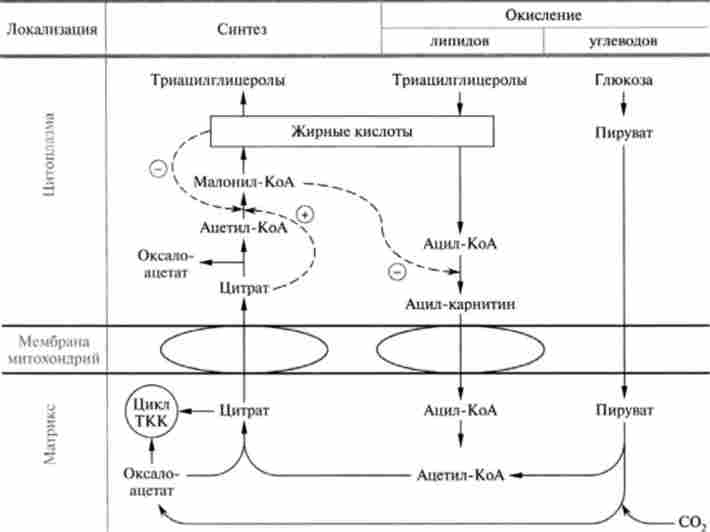
Мал. 23.18. Схема регуляції окислення і синтезу жирних кислот: пунктиром показані позитивні (+) і негативні (-) ефекти карбоксилазу, що призводить до синтезу Малоні-КоА, а отже, ініціюється синтез жирних кислот. У свою чергу, Малоні-КоА інгібує кар- Нітін-ацілтрансферази - фермент, що забезпечує транспорт ацетил-КоА в матрикс мітохондрії, де відбувається його окислення. Отже, активація біосинтезу жирних кислот автоматично вимикає їх розпад. Якщо ж концентрація глюкози знижується, а відповідно зменшується концентрація і оксалоацетата, створюються умови, що відкривають шлях для жирних кислот в мітохондрії, де починається їх окислення, що забезпечує потреби клітини в енергії.
Особливу роль в регуляції метаболізму ліпідів грають гормони. Слід звернути увагу на те, що жировий обмін регулюється практично тими ж гормонами, що і обмін вуглеводів - адреналін і норадреналін, Глюка гон, глюкокортикоїди, гормони передньої долі гіпофіза (соматотропний гормон і АКТГ), а також тироксин і статеві гормони. Адреналін і норадреналін активують ліполіз в жировій тканині, в результаті посилюється мобілізація жирних кислот з жирових депо і зміст неестеріфіцірованних жирних кислот в плазмі підвищується. Як вже зазначалося (гл. 23.3), ці гормони через цАМФ активують відповідну протеїн, яка сприяє фосфорилированию ліпази, т. Е. Утворення її активної форми.
Крім цього, відомо, що глюкагон і адреналін через цАМФ-зависи- мую протеинкиназу каталізують фосфорилювання ацетил-КоА-карбокси- лази і переводять її в неактивну форму, тим самим пригнічуючи процеси ли погенеза. Що стосується механізму регуляторного дії СТГ і АКТГ, також активують процеси ліполізу, то первинний механізм їх дії пов'язаний, мабуть, з індукцією синтезу аденілатциклази і гормончувстві- котельної ліпази. Тут доречно нагадати, що якщо адреналін стимулює ліполіз майже миттєво, то дія гормонів гіпофіза на ліполіз характеризується наявністю досить тривалої лаг-фази.
Інсулін надає протилежне адреналіну і глюкагону дію на ліполіз і мобілізацію жирних кислот. В даний час встановлено, що інсулін стимулює фосфодіестеразную активність в жировій тканині і таким чином грає важливу роль в підтримці стаціонарного рівня цАМФ в тканинах, а отже, і утворення активної форми ліпази. Інсулін надає стимулюючу дію на процеси біосинтезу жирних кислот і триацилгліцеролів, окислення глюкози і утворення пірувату. Всі ці ефекти залежать від концентрації глюкози і можуть бути пояснені здатністю інсуліну збільшувати надходження глюкози в клітини жирової тканини.
Інші гормони, зокрема гормони щитовидної залози, статеві гормони, самі по собі не мають прямого впливу на ліполіз, діють опосередковано, найчастіше як фактори, що стимулюють дію інших гормонів.