| Головна |
| «« | ЗМІСТ | »» |
|---|
Периферична (рецепторна) частина систем слуху і рівноваги (вестибулярного аналізатора) мають загальні риси. Рецептори цих двох систем є волосовими клітинами, т. Е. Механорецепторами, і розташовуються в структурах внутрішнього вуха. Внутрішнє вухо, в свою чергу, є продовженням зовнішнього і середнього вуха і тісно пов'язане з ними анатомічно і функціонально.
Будова зовнішнього, середнього і внутрішнього вуха. Навколишній світ сповнений звуків - акустичних коливань повітря. Як відомо, людина чує звуки в діапазоні від 16 до 20 000 Гц, що помітно вже, ніж у багатьох тварин (дельфінів, собак, кішок), і пов'язано зі специфікою мовних, т. Е. Комунікативних, сигналів, притаманних нашому біологічному виду. Слухова система включає зовнішнє, середнє і внутрішнє вухо.
Зовнішнє вухо складається з вушної раковини і зовнішнього слухового проходу. Форма і рельєф вушної раковини грають значну роль в сприйнятті звуків. Якщо цей рельєф змінити, наприклад затоку воском, людина буде гірше визначати напрямок на джерело звуку. Зовнішнє вухо відділене від середнього барабанною перетинкою, що має вигляд конуса з вершиною, зверненої в бік равлики.
Середнє вухо людини складається з барабанної порожнини, з'єднаної евстахиевой трубою з порожниною глотки. Крім барабанної перетинки, до нього відносять зчленовані між собою три слухові кісточки: молоточок, ковадло і стремечко. Євстахієву труба служить для вирівнювання тиску повітря по обидві сторони барабанної перетинки (між зовнішнім слуховим проходом і барабанної порожниною). Слухові кісточки діють як важелі, забезпечуючи передачу звукових коливань на овальне вікно (область контакту стремечка і равлики). Енергія звуку, зібрана з відносно великої площі барабанної перетинки, через слухові кісточки передається на малу площу овального вікна. В результаті відбувається зростання тиску, достатній для того, щоб коливання середовища з низькою щільністю (повітря) приводили до коливань ендолімфи усередині равлики. Система слухових кісточок включає в себе також дві маленькі м'язи, одна з яких прикріплена до ручки молоточка, а інша - до стременця. Скорочувальна активність цих м'язів запобігає занадто великі коливання, викликані гучними звуками. Ця реакція називається акустичним рефлексом, основна функція якого полягає в захисті внутрішнього вуха від ушкоджує звуковий стимуляції.
внутрішнє вухо включає два рецепторних апарату: слуховий і вестибулярний (рис. 12.3).
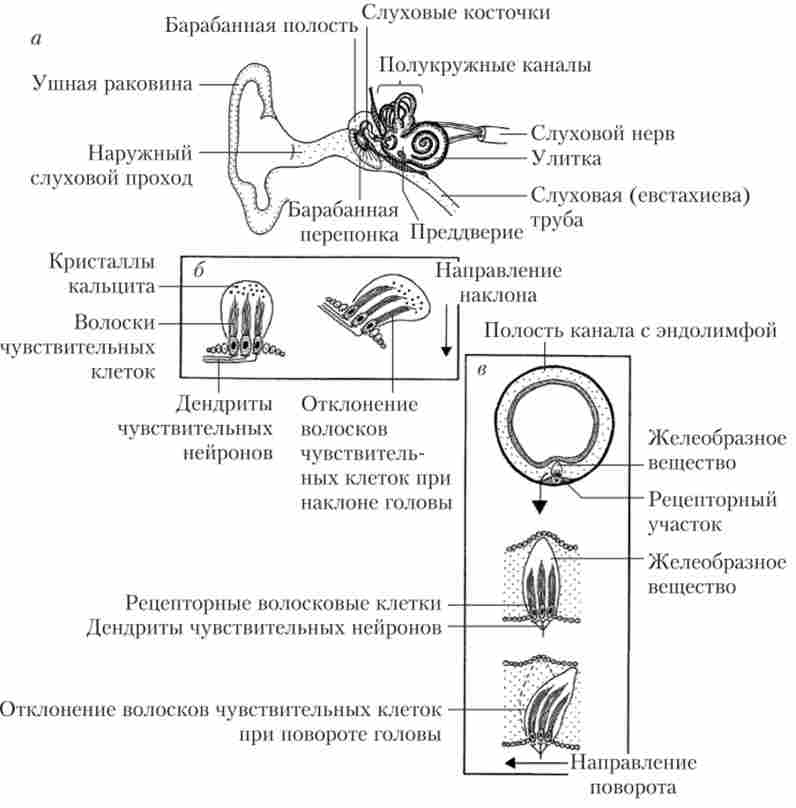
Мал. 123. Будова слухового і вестибулярного апаратів вуха:
а - основні структури вуха; б - рецептори передодня; в - рецептори півколових каналів
Слуховий рецепторний апарат розташовується в структурах равлики, вестибулярний - в структурах вестибулярних мішечків і півколових каналів. Всі частини системи внутрішнього вуха лежать в особливих порожнинах скроневих кісток.
Улітку розташовується в кістковому каналі, розділеному двома мембранами - базилярної, або основний, і рейснеровой, або вестибулярної, - на три окремих каналу (сходи): барабанний, середній і вестибулярний (рис. 12.4).
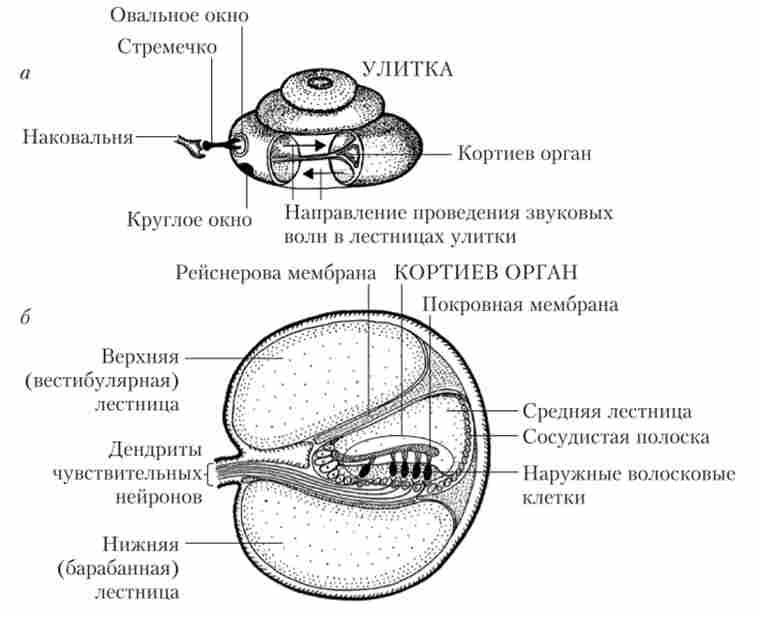
Мал. 12.4. Внутрішньо будова органу слуху:
а - основні елементи равлики; б - сходи равлики і будова кортиева органу
Сходи внутрішнього вуха заповнені рідинами, іонний склад яких специфічний в кожному каналі. Середня сходи заповнена ендолімфою з високим вмістом іонів К+; вона повідомляється з ендолімфою вестибулярних органів. Дві інші сходи заповнені перілім- фої, за складом подібна до тканинної рідиною (з низьким вмістом іонів К+). Усередині середньої сходи є кортиев тунель. По обидва боки від нього знаходяться опорні клітини і ряди волоскових клітин (слухові рецептори), звані відповідно зовнішніми і внутрішніми волосовими клітинами. Волоскові і опорні клітини, а також кортиев тунель покриті текторіальная (покривної) сполучнотканинною мембраною. Рухи текторіальная мембрани відіграють важливу роль в активації слухових рецепторів і розвитку на них генераторного потенціалу. Описана структура, що тягнеться по всій довжині равлики, носить назву «кортів орган» (рис. 12.4, б). Барабанна і вестибулярна сходи на вершині равлики з'єднуються один з одним через невеликий отвір - гелікотрему, а середня сходи - сліпо замкнута.
Вестибулярних мішечків всього два: один з них називається утрікулюс (маточка), а другий - саккулюс (власне мішечок). Разом вони утворюють переддень равлики. Зсередини на одній зі стінок кожного мішечка знаходяться волоскові (рецепторні) клітини. Мішечки пов'язані один з одним, а утрікулюс, крім того, з'єднаний з напівкружними каналами.
Три напівкружних каналу розташовані в площинах, що утворюють тривимірну систему координат (взаємно перпендикулярні). Кожен канал виходить з утрікулюса і, описавши близьку до півкола дугу, знову приходить до нього з іншого боку. Близько утрікулюса канал розширюється приблизно в 10 разів, утворюючи ампулу. В ампулі знаходяться сенсорні клітини, волоски яких пронизують канальці желеподібної маси - купули. Освіти, аналогічні кунуле, є і в вестибулярних мішечках.
Волоскові клітини внутрішнього вуха є вторинно чувству- ющими рецепторами, т. е. не мають аксона і не є нейронами. Зовнішній механічний стимул викликає генерацію волоскової кліткою рецепторного потенціалу, що забезпечує секрецію медіатора (глутамінової кислоти) в синаптичну щілину. Це призводить до виникнення на мембрані аферентного волокна генераторного потенціалу, а в разі перевищення порога - імпульсного відповіді (потенціалів дії). У нижній частині рецепторной клітини локалізуються як аферентні, так і еферентні синапси. Завдяки такій іннервації чутливістю рецептора може управляти ГМ. Апікальна (верхня) частина волоскової клітини знаходиться в ендолімфі, в разі слухового рецептора від неї відходять 30-150 виростів, або стереоцилій, впорядковано розташованих на поверхні клітини. У разі вестибулярних рецепторів в пучок волосків включена одна велика кіноцілій, що представляє собою видозмінену нерухому вію.
Відповідь волоскової клітини визначається зміною іонної провідності апікальної мембрани. У спокої клітина має невелике число відкритих К ~ -каналів, через які входять іони К+. Зсув пучка стереоцилій в сторону кіноцілій збільшує число відкритих калієвих каналів і деполяризує рецептор, зміщення в протилежний бік (від кіноцілій) - гіперполяризуючий. За рахунок закриття калієвих каналів порогова величина зміщення волосків становить 10-100 нм. На мікроскопічному рівні показано, що зі стулками механочувствітельпих калієвих каналів пов'язані найтонші білкові нитки, що йдуть від кіноцілій до стереоцилиями, а також від однієї стереоціліямі до іншої.
Особливості роботи слуховий системи. Розрізнення частот. Звукова хвиля тисне на барабанну перетинку. Це тиск через систему слухових кісточок передається на овальне вікно равлики і впливає на перилімфу вестибулярної сходи. Так як перилимфа нестислива, коливання звукових хвиль передаються по ній через гелікотрему в барабанну драбину, а звідти - на базилярную мембрану. Коливання базилярної мембрани ведуть до зміщення стереоцилій волоскових клітин по відношенню до текторіальная мембрані, і, отже - до деполяризації або гіперполяризації мембрани волоскових клітин (і далі - до реакції волокон слухового нерва).
Будова і властивості волокон базилярної мембрани такі, що різні її частини збільшують амплітуду своїх коливань (входять в резонанс) при дії звукової хвилі тільки певної частоти. При цьому волоскові клітини, розташовані в різних ділянках базилярної мембрани, «інформують» ГМ про наявність в звуці певних тональностей. Широка частина равлики (далеко від овального вікна) і її волоскові клітини реагують на високі частоти. На звуки низької частоти реакцію забезпечують базилярная мембрана і рецепторні клітини в вузької частини равлики.
Таким чином, в равлику протікає частотно-амплітудний аналіз звукових хвиль, і в ЦНС по слуховому нерву відправляється інформація про те, скільки звукових коливань кожної частоти впливає на організм в даний момент.
Слухові центри головного мозку. Потрапивши по слуховому нерву в задній мозок, на першому етапі сигнали, що йдуть від волоскових клітин, аналізуються нейронами довгастого мозку і моста. Основна мета такого первинного аналізу полягає в порівнянні сигналів від правої і лівої равликів, яке допомагає визначити напрям на джерело звуку. Після цього слухова інформація відправляється в нижні горбки четверохолмия (середній мозок). Подібно згадуваним раніше верхнім горбка, нейрони в цій області реагують на нові звукові сигнали з наступним запуском орієнтовного рефлексу. Він проявляється в повороті очей і голови в бік джерела звуку.
Наступний слуховий центр знаходиться в медіальному колінчастому тілі таламуса. Тут при перемиканні відбувається фільтрація сигналів перед їх надходженням в кору великих півкуль. Таламус «пригнічує» в потоці проводиться слуховий інформації зайві шуми, забезпечує «слухове увагу» (налаштування слуховий системи на сприйняття частот певного діапазону). Так, при звучанні двох розрізняються голосів одночасно (наприклад, чоловічого і жіночого) ми відносно легко налаштовуємося на сприйняття одного з них.
слухова (скронева) кора, як і зорова, підрозділяється на первинну, вторинну і третинну зони.
Первинна слухова кора розташована в самій верхній частині скроневої частки. Тут завершується частотно-амплітудний аналіз. Діапазони тональностей, на сприйняття яких налаштовані нейрони в первинній слуховий корі, дуже вузькі (2-3 Гц).
Вторинна слухова кора лежить нижче первинної. Вона відповідає за сприйняття нескладних слухових образів, що представляють собою суму декількох тональностей. До таких слуховим образам відносяться різноманітні шуми, скрипи і шерехи ( «звуки природи»), а також видоспецифічні сигнали, що видаються людиною, - плач, сміх, крик болю.
Третинна слухова кора розташовується переважно в задній області скроневої частки. Дана частина кори великих півкуль забезпечує розпізнавання найбільш складних звукових образів, значущим ознакою яких є не сума, а співвідношення тональностей (музика і мова).
Дійсно, мелодія пізнавана незалежно від того, зіграна вона на скрипці або на віолончелі; слово впізнається незалежно від того, який голос його вимовив (чоловічий, жіночий або дитячий). Чоловічі голоси звучать здебільшого на частотах близько 100 Гц, жіночі - в районі 200 Гц, а дитячі - близько 400 Гц (дворазове збільшення частоти відповідає переходу з однієї октави в іншу).
Частина третинної слуховий кори, що відповідає за сприйняття слів, отримала власне назва - зона Верніке. Ушкодження цієї області веде до нездатності розуміти мову (мовна агнозия). При цьому людина зазвичай продовжує розуміти написані слова. Аксонинейронів зони Верніке йдуть в речедвигательную область кори великих півкуль - зону Брока. Таким чином, дві дані структури виявляються анатомічно пов'язаними. Це призводить до того, що при ураженні зони Верніке страждає і експресивна мова (втрачається здатність підібрати потрібне слово і вимовити його). Показано, що пацієнти з ураженнями верхніх відділів скроневої ділянки мозку здатні розрізняти прості звуки, їх можна легко навчити розрізняти топи по висоті, але вони не здатні розрізняти і відтворювати групи ритмічних ударів в досить швидкому темпі.
Особливості роботи вестибулярної системи. Найбільш загальне призначення вестибулярної системи полягає в реакції на прискорення. Сидячи, наприклад, в кріслі поїзда, ми навіть із закритими очима відчуваємо, як він гальмує, набирає швидкість, повертає. Виникнення цих відчуттів і становить функцію вестибулярного аналізатора.
При появі прискорення відбувається інерційне зміщення желеподібних мас, що лежать в вестибулярних мішечках і півколових каналах, щодо стінок внутрішнього вуха. Це призводить до вигину стереоцилій рецепторних клітин і спрацьовування рецепторів. Різні рецептори налаштовані на різні напрямки прискорення, оскільки ряди ( «гребені») їх волосків орієнтовані в просторі по-різному.
Рецептори вестибулярних мішечків реагують на лінійні прискорення (зміщення вперед-назад, вправо-вліво і т. П.). Рецептори півколових каналів відповідають на кутові прискорення (повороти і обертання) (рис. 12.3, б, в).
На волоскові клітини ампул півколових каналів впливає переміщення навколишнього їх желеподібної маси купул. Волоски цих рецепторних клітин мають відносно велику довжину. На коротші волоски рецепторів вестибулярних мішечків впливають руху аналогічної купуле отолитовой мембрани - покриває волоски желеподібної маси, обтяженою (в порівнянні з ендолімфою) Отоліти, т. е. кристалами кальциту (СаС03) Розміром до 10 мкм. Отоліто- вая мембрана пронизана впорядкованими стереоцилиями рецепторних волоскових клітин, які разом з підтримуючими клітинами утворюють сенсорний епітелій - макулу. При лінійному прискоренні отолитовой мембрана завдяки інерції ковзає по макуле і зрушує стереоціліямі волоскових клітин.
Один з видів лінійного прискорення - прискорення вільного падіння ( «g», від gravitation - гравітація). Воно діє на організм постійно.
Відповідно, частина рецепторів мішечків завжди активована. Завдяки цьому ми отримуємо інформацію про положення голови в просторі, після чого наш мозок може визначити положення в просторі всього тіла (з урахуванням вигинів хребта і суглобів кінцівок).
Вестибулярна інформація надзвичайно важлива для управління рухами. У зв'язку з цим, потрапивши через вестибуло-слуховий нерв в задній мозок, вестибулярні сигнали швидко поширюються по ЦНС. Вони необхідні: