| Головна |
| «« | ЗМІСТ | »» |
|---|
Вміщені в клітці РНК розрізняються складом, розміром, функціями і локалізацією.
У цитоплазмі клітин містяться три основних функціональних виду РНК. це матричні РНК (мРНК), що виконують функції матриць білкового синтезу, Хвороби РНК (рРНК), що виконують роль структурних компонентів рибосом, і транспортні РНК (тРНК), які беруть участь в трансляції (перекладі) інформації мРНК в послідовність амінокислот у білку.
В ядрі клітин виявляють ядерну РНК, що становить від 4 до 10% ог сумарною клітинної РНК. Основна маса ядерної РНК представлена високомолекулярними попередниками рибосомних і транспортних РНК. Попередники високомолекулярних рРНК (28 S, 18 S і 5 S РНК) в основному локалізуються в полісом. Від 2 до 10% від сумарної ядерної РНК припадає на особливу фракцію гетерогенної ядерної РНК (г-яРНК), молекули якої є попередниками мРНК.
Важливим функціональним типом ядерної РНК є малі ядерні РНК (м-яРНК), що містять від 90 до 300 нуклеотидів з унікальною нуклеотидної послідовністю, комплементарної послідовністю сайтів сплайсингу. Завдяки цьому м-яРНК регулює дозрівання (процесинг) r-яРНК в зрілі мРНК.
РНК є основним генетичним матеріалом у деяких вірусів тварин і рослин (геномні РНК). Для більшості РНК вірусів характерна зворотна транскрипція їх РНК геному, яку направляють зворотного транскриптазой.
Все РНК являють собою полімери рібонуклс- відійшов, з'єднаних, як в молекулі ДНК. 35'-фосфорнодіефірнимі зв'язками. На відміну від ДНК, що має двухцепочечную структуру, РНК являє собою одноцепочечниє лінійні полімерні молекули.
До теперішнього часу вдалося визначити первинну структуру більшості тРНК, рРНК, мРНК і м-яРНК з різних видів живих організмів і виявити основні закономірності їх структурної організації.
Структурна організація мРНК. мРНК - найбільш гетерогенний щодо розмірів і стабільності клас РНК. Зміст мРНК в клітинах становить 2-6% від тотального кількості РНК. мРНК, особливо еукаріотичні, мають деякими специфічними структурними особливостями. мРНК складаються з ділянок - ціетронов, що визначають послідовність амінокислот в кодованих ними білках, і нетрансльовані областей на кінцях молекули. Для цистрона областей характерна унікальна послідовно ність нуклеотидів, що визначається нуклеотидної послідовністю гена, нетрансльовані області мають деякі загальні закономірності нуклеотидного складу будови. Так, на 5'-кінці всіх еукаріотичних мРНК є особлива структура, звана кепом (Від англ, cap- ковпачок). Кеп є 7-метілгуанозінтріфосфат, приєднаний до 5'-гідро- ксиліт кінцевого, як правило, 2-о-мстілрібонуклсозіда через залишок три- фосфату. Освіта кепа відбувається ферментативним шляхом в ядрі ще до завершення транскрипції. Вважається, що кеп, з одного боку, оберігає 5'-кінець мРНК від її розщеплення 5'-екзонуклсазамі, з іншого боку, використовується для специфічного впізнавання в системі трансляції. За кепом слід прецістронний нетрансльовані ділянку, в якому (від 3-15 нуклеотидів до ініціюючого кодону) розташовується послідовність нуклеотидів, комплементарная послідовність рРНК. Її роль - забезпечення правильного взаємодії 5'-кінця з рибосомою. Завершується цистрон терминирующего кодоном, за яким слід постцістронний нетрансльовані ділянку, що має в своєму складі характерний для багатьох видів гексануклеотід ААУААА. У більшості мРНК З'-кінець містить поліадені- латну ланцюжок з 20-250 аденілових нуклеотидів, що не є результатом транскрипції, а приєднуються до мРНК в ході дозрівання в ядрі ферментативним шляхом. Передбачається, що поліадснплатная послідовність відповідає за підтримання внутрішньоклітинної стабільності мРНК, визначає її час існування.
мРНК володіють складною вторинної структурою, що забезпечує виконання ними матричної функції в ході трансляції. Показано, що в цілому в лінійної молекули мРНК формується кілька двухспіральной шпильок, на кінцях яких розташовуються «знаки» ініціації і термінації трансляції.
Структурна організація тРНК. Транспортні РНК виконують функції посередників (адаптерів) в ході трансляції мРНК. Кожній з 20 протеіногенних амінокислот відповідає своя тРНК. Для деяких амінокислот, які кодуються двома і більше кодонами, існують кілька тРНК.
тРНК являють собою порівняно невеликі одноцепочечниє молекули, що складаються з 70-93 нуклеотидів. Їх молекулярна маса становить
(2,4-3,1) - 104kDa. На долютРНК припадає приблизно 15% сумарної клітинної РНК.
До теперішнього часу встановлено нуклеотидних послідовність майже для 300 тРНК, виділених з різних видів організмів і володіють різною амінокислотної специфічністю. Незважаючи на відмінності в нуклеотидної послідовності, все тРНК мають багато спільних рис. У всіх тРНК вісім або більше нуклеотидів містять різні мінорні модифіковані підстави (всього близько 60), багато з яких представляють собою метиловані пуринові або піримідинові підстави. Обов'язковими мінорними компонентами для всіх тРНК є дігідроурідін і псевдоуридин. У більшості тРНК на 5'-кінці знаходиться залишок гуаніло- вої кислоти, а на -кінці всіх тРНК, званому акцепторним, обов'язковим є трінуклеотідамі-Ц-Ц-А (3 ').
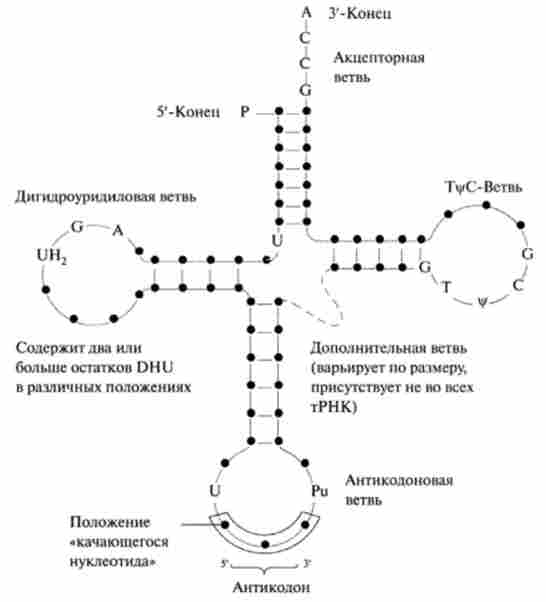
Вторинна структура тРНК формується за рахунок утворення максимального числа водневих зв'язків між внутрішньомолекулярними комплементарними парами азотистих основ. В результаті утворення цих зв'язків поли нуклеотидная ланцюг тРНК закручується з утворенням спіраль гілок, що закінчуються петлями з неспарених нуклеотидів. Просторове зображення вторинних структур всіх тРНК має форму листа конюшини (рис. 14.7).
Мал. 14.7. Вторинна структура тРНК
Мал. 14.8. Третинна структура тРНК (по А. С. Спіріну)
У «лист конюшини» розрізняють чотири обов'язкові гілки, довші тРНК, крім того, містять коротку п'яту (додаткову) гілка.
Адапторная функцію тРНК забезпечують акцепторная гілка, до -кінцю якої приєднується ефірним зв'язком амінокислотний залишок, і протистоїть акцепторной гілки антикодоновая гілка, на вершині якої знаходиться петля, що містить антикодон. Антикодон є специфічним триплет нуклеотидів, який комплементарний в антіпарал- лсльном напрямку кодону мРНК, що кодує відповідну амінокислоту.
Т-Гілка, несуча петлю псевдоуридин (Т | / С-петлю), забезпечує взаємодію тРНК з рибосомами. Л-гілка, несуча дегідроурідіновую петлю, найімовірніше забезпечує взаємодію тРНК з відповідною аміноацил-тРНК-синтетазой. Функції п'ятої додаткової гілки поки мало досліджені, найімовірніше вона зрівнює довжину різних молекул тРНК.
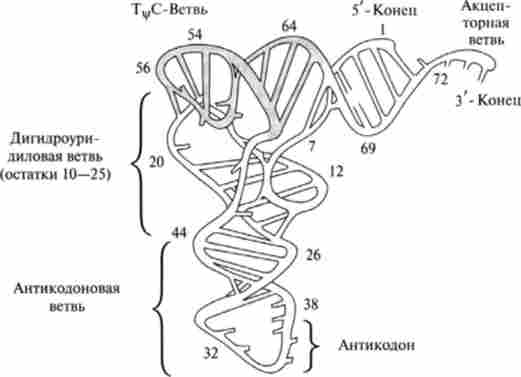
Третинна структура тРНК дуже компактна і утворюється шляхом зближення окремих гілок конюшини за рахунок додаткових водневих зв'язків і стекінг-взаімодсйствій з утворенням L-образної структури «ліктьового згину» (рис. 14.8). При цьому акцепторна плече, що зв'язує амінокислоту, виявляється розташованим на одному кінці молекули, а антикодон - на іншому. Третинні структури всіх тРНК настільки схожі, що суміш різних тРНК утворює кристали. У той же час наявні в просторову структуру незначні відмінності забезпечують специфічне впізнавання тРНК відповідними аміноацил-тРНК-синтетазами.
Структура Хвороби і рибосом. Хвороби формують ту основу, з якою зв'язуються специфічні білки при утворенні рибосом. Рибосоми - це нуклеопротеіновие органели, що забезпечують синтез білка на мРНК-матриці. Число рибосом в клітці дуже велике: від 104 у прокаріотів до 106 у еукаріот. Локалізуються рибосоми головним чином в цитоплазмі, у еукаріот, крім того, в полісом, в матриксі мітохондрій і стромі хлоронластов. Рибосоми складаються з двох субчастиц: великої і малої. За розмірами і молекулярної масі все вивчені досі рибосоми ділять на 3 групи - 70S рибосоми прокаріотів ^ що складаються з малої 30S і великий 50S субчастиц, 80S рибосоми еукаріот, що складаються з 40S малої і 60S великий субчастиц, і рибосоми мітохондрій і хлоронластов, які в обшем відносять до класу 70S, проте вони розрізняються але коефіцієнтами седиментації у різних груп еукаріот.
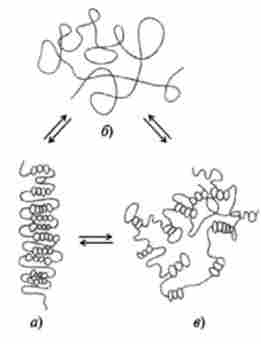
Мал. 14.9. Третинна структура Хвороби в розчині в залежності від іонної сили, температури і pH середовища (по А. С. Спіріну): а - компактна паличка; б - розгорнута ланцюг; в - компактний клубок
Мала субчастіца 80S рибосом утворена однією молекулою рРНК (18S) та 33 молекулами різних білків. Велика субчастіца утворена трьома молекулами рРНК (5S, 5,8S і 28S) та приблизно 50 білками. Все рРНК, за винятком 5S РНК, мають спільного попередника - 45S РНК, локалізовану в полісом.
Прокаріотичні рибосоми і рибосоми мітохондрій і пластид містять менше компонентів, але структурно і функціонально дуже схожі з еукариотичними. Вторинна структура рРНК утворюється за рахунок коротких двоспіральної ділянок молекули - шпильок (рис. 14.9). Близько 2/3 рРНК організовано в шпильки, 1/3 - представлена однотяжевимі ділянками, багатими пуриновими нуклеотидами, з якими переважно зв'язуються білки. Білки рибосом, подібно гістонів, мають основним характером, виконують як структурну, так і ферментативну роль.
Дослідження останніх років показали, що Хвороби є не тільки структурними компонентами рибосом, але і забезпечують правильне зв'язування їх з певною нуклеотидної послідовністю мРНК, встановлюючи тим самим початок і рамку зчитування при утворенні поліпеп- тідной ланцюга. Крім того, рРНК беруть участь в забезпеченні взаємодії рибосом з тРНК.
В рибосомах є дві борозенки, одна з них утримує мРНК, інша - зростаючу поліпептидний ланцюг. Крім цього, в рибосомах є дві ділянки, що зв'язують тРНК-аміноацільний (A-ділянка) і пептіділь- ний (П-ділянка). Освіта і функціонування А- і П-ділянок забезпечується обома субчастиц рибосом.