| Головна |
| «« | ЗМІСТ | »» |
|---|
Одна з особливостей живих організмів полягає в тому, що всі вони являють собою відкриті системи, які здатні витягувати, перетворювати і використовувати енергію навколишнього середовища або в формі органічних поживних речовин (хемотрофи), або у формі енергії сонячного випромінювання (фототрофи). Обмін енергією в організмі тісно пов'язаний з обміном речовин (метаболізмом). Метаболізм можна визначити як сукупність ферментативних хімічних реакцій, які можуть протікати в клітці. Активність ферментів, які каталізують ці реакції, регулюється за допомогою чутливої системи взаємопов'язаних механізмів, тому метаболізм являє собою висококоорлінірованную, цілеспрямовану клітинну активність. Він виконує наступні функції:
Перетворення органічних сполук у клітині здійснюється, як правило, у вигляді ланцюга або послідовності реакцій, які називаються метаболічними шляхами. а залучаємо в такі реакції з'єднання - метаболітами. У класичній біохімії метаболічні шляхи поділяються на два типи: катаболічні і анаболічні. Катаболічсскіс шляху - це процеси ферментативної деградації, в ході яких великі органічні молекули руйнуються (зазвичай в окислювальних реакціях) до простих клітинних компонентів з одночасним виділенням вільної хімічної енергії. Ця енергія використовується потім організмом для підтримки життєдіяльності, зростання і реплікації, а також перетворюється в інші форми енергії - механічну, електричну і теплову.
Анаболічні шляхи - це процеси ферментативного синтезу, в ході яких з відносно простих попередників будуються складні органічні компоненти клітини; синтез часто включає відновлювальні етапи і супроводжується витратою вільної хімічної енергії (рис. 15.1).
Всі метаболічні системи відрізняються впорядкованістю і простотою, не дивлячись на різноманітність метаболітів, як споживаних, так і утворюються. Особливо важливе значення має відкриття центральних шляхів обміну, які примикають і до катаболичним, і до анаболічних шляхах, т. е. безпосередньо пов'язують між собою ге та інші.
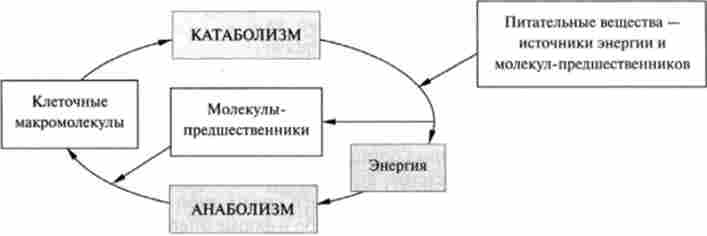
Мал. 15.1. Взаємозв'язок між катаболичними і анаболічними процесами
Таким чином, обмін речовин тісно пов'язаний з обміном енергії. Реакції катаболізму, що супроводжуються зменшенням вільної енергії (-ДСг), є донорами не тільки структурних попередників, а й забезпечують енергетично процеси анаболізму (+ Д (7). Нагадаємо, що якщо Д (7 негативно, то реакція протікає мимовільно і супроводжується зменшенням вільної енергії . Такі реакції називаються екзергонічнолш, до них відносяться, як правило, катаболічні перетворення. Якщо ж значення ДG позитивно, то реакції будуть протікати тільки при надходженні вільної енергії ззовні і називатися ендергонічеськие (Анаболічні процеси). При Д (/, що дорівнює нулю, система знаходиться в рівновазі.
Слід зазначити, що зміна вільної енергії, певне при стандартних умовах (концентрація реагентів 1 моль / л, pH 0), називається зміною вільної стандартної енергії і позначається ДСГ. при визначенні AG0 в біохімічних реакціях, для яких стандартною умовою є значення pH, рівне 7,0, ця величина позначається як Д<Г
Як відомо, в біоенергетиці живих організмів мають значення два основних моменти:
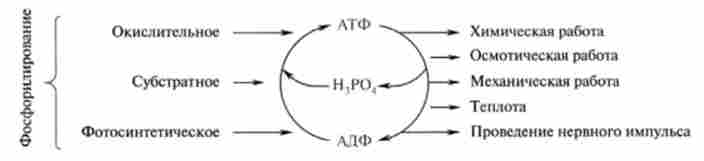
Мал. 15.2. Обмін АТФ в клітинної енергетики: справа - процеси, що вимагають витрати енергії; зліва - типи синтезу АТФ в природі шляхомфосфорилювання АДФ
Постає питання, чому молекула АТФ відповідає своїй центральній ролі в біоенергетиці. Для його вирішення розглянемо структуру АТФ:
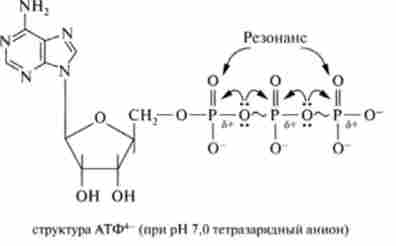
АТФ є термодинамічно нестійке з'єднання. Нестабільність АТФ визначається, по-перше, електростатичним Отта / шіва- ням в області кластера однойменних негативних зарядів, що призводить до напруження всієї молекули, однак найсильніше зв'язку - Р-О-Р, і, по-друге, конкурентним резонансом. Відповідно до останнього фактором існує конкуренція між атомами фосфору за неподіленого рухливі електрони атома кисню, розташованого між ними, оскільки на кожному атомі фосфору є частковий позитивний заряд внаслідок значного елсктронакцепторного впливу груп Р = 0 і Р-О-. Таким чином, можливість існування АТФ визначається наявністю достатньої кількості хімічної енергії в молекулі, що дозволяє компенсувати ці фізико-хімічні напруги. У молекулі АТФ є дві фосфоан- гідридних (пірофосфатних) зв'язку, гідроліз яких супроводжується значним зменшенням вільної енергії (при pH 7,0 і 37 ° С):
Зв'язки, при гідролізі яких зміни вільної енергії системи складають понад 30 кДж / моль, в біохімії називають макроергічними і позначають знаком ~ (тильда), а сполуки, які мають такими зв'язками, - макроергів.
До сполукам, які мають макроергічними зв'язками, крім АТФ, відносяться також УТФ, ЦТФ, ГТФ, ТТФ, крсатінфосфат, деякі тіоефі- ри (наприклад, ацил-КоА), фосфоснол бенкеті ват, 1,3-діфосфагліцерат і деякі інші сполуки. Однак утворення цих макроергічсскіх з'єднань в більшості випадків залежить від енергії, що поставляється АТФ.
Однією з центральних проблем біоенергетики є біосинтез АТФ, який в живій природі відбувається шляхом фосфорилювання АДФ.
Фосфорилювання АДФ є ендергонічеськие процесом і вимагає джерела енергії. Як зазначалося раніше, в природі переважають два таких джерела енергії - це сонячна енергія і хімічна енергія відновлених органічних сполук.
Зелені рослини і деякі мікроорганізми здатні трансформувати енергію поглинених квантів світла в хімічну енергію, яка витрачається на фосфорилювання АДФ в світловий стадії фотосинтезу. Цей процес регенерації АТФ отримав назву фотосинтетичного фосфорші- вання (Гл. 16).
Трансформація енергії окислення органічних сполук в макроер- ня зв'язку АТФ в аеробних умовах відбувається переважно шляхом окисного фосфорилювання. Вільна енергія, необхідна для утворення АТФ, генерується вдихательная окисної ланцюга мітохондрій (15.3.2).
Відомий ще один тип синтезу АТФ, який отримав назву субстратного фосфорилювання. На відміну від окисного фосфорилювання, сполученого з перенесенням електронів, донором активованої фосфорильної групи (~ Р03Н2), Необхідної для регенерації АТФ, є інтермедіантов процесів гліколізу (гл. 18) і циклу трикарбонових кислот (гл. 19). У всіх цих випадках окислювальні процеси призводять до утворення високоенергетичних сполук: 1,3-дифосфоглицерата (гліколіз), сукцинил-КоА (цикл трикарбонових кислот), які за участю відповідних ферментів здатні фосфоріліровагь АДФ і утворювати АТФ. Трансформація енергії на рівні субстрату є єдиним шляхом синтезу АТФ в анаеробних організмах. Цей процес синтезу АТФ дозволяє підтримувати інтенсивну роботу скелетних м'язів в періоди кисневого голодування. Слід пам'ятати, що він є єдиним шляхом синтезу АТФ в зрілих еритроцитах, які не мають мітохондрій.