| Головна |
| «« | ЗМІСТ |
|---|
По двох головних подій митотичного циклу в ньому виділяють репродуктивну і розділову фази, відповідні інтер фазетл митозу класичної цитології (рис. 2.11).
У початковий відрізок інтерфази (постмітотічних, пресинтетичний, або G ^ -nepuod) відновлюються риси організації інтерфазних клітини, завершується формування ядерця, що почалося ще в телофазе. З цитоплазми в ядро надходить значна (до 90%) кількість білка. У цитоплазмі паралельно реорганізації ультраструктури інтенсифікується синтез білка. Це сприяє зростанню маси клітини. Якщо дочірній клітині належить вступити в наступний мітотичний цикл, синтези набувають спрямований характер: утворюються хімічні попередники ДНК, ферменти, що каталізують реакцію редуплікаціі ДНК, синтезується білок, початківець цю реакцію. Таким чином здійснюються процеси підготовки наступного періоду інтерфази - синтетичного.
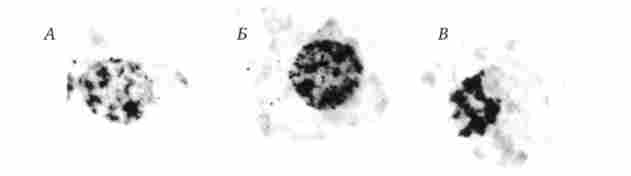
Мал. 2.11. Інтерфазна і мітотична форми структурної організації хромосом.
А - хроматин в інтерфазних ядрі; Б - включення в ядро радіоактивного попередника ДНК в синтетичному періоді мітотичного циклу; В - клітина в анафазе мітозу
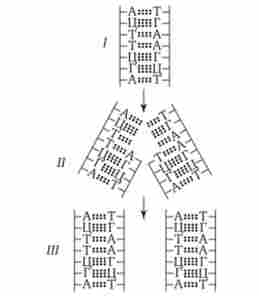
Мал. 2.12.
Напівконсервативний спосіб редуплікаціі ДНК.
/ - материнська біспіралью ДНК; II - добудовування комплементарних полінук- леотідних ланцюгів; III - дві дочірні биспирали ДНК; А, Т, Г, Ц - символи азотистих основ нуклеотидів
В синтетичному, або S-періоді, подвоюється кількість спадкового матеріалу клітини. За малими винятками редупликация ДНК здійснюється напів консервативним способом (рис. 2.12). Він полягає в розходженні биспирали ДНК на два ланцюги з подальшим синтезом біля кожної з них комплементарної ланцюжка. В результаті виникають дві ідентичні биспирали (деталі див. Розд. 3.4.2.1). Молекули ДНК, комплементарні материнським, утворюються окремими фрагментами по довжині хромосоми, причому неодномоментно (асинхронно) в різних ділянках однієї хромосоми, а також в різних хромосомах. Потім ділянки (одиниці реплікації - Реплі- кони) Новоствореної ДНК «зшиваються» в одну макромолекулу.
У клітці людини міститься більше 50 000 репліконов. Довжина кожного з них близько 30 мкм. Число їх змінюється в онтогенезі. Сенс редуплікаціі ДНК РЕПЛІКОН стає зрозумілим з наступних зіставлень. Швидкість синтезу ДНК становить 0,5 мкм / хв. В цьому випадку редупликация нитки ДНК однієї хромосоми людини довжиною близько 7 см повинна була б зайняти близько трьох місяців. Тривалість же синтетичного періоду в клітинах людини становить 7-12 год.
Ділянки хромосом, в яких починається синтез, називають точками ініціації. Можливо, ними є місця прикріплення інтерфазних хромосом до внутрішньої мембрані ядерної оболонки. У клітці, що пройшла S-період інтерфази, хромосоми містять подвійну кількість генетичного матеріалу. Поряд з ДНК в синтетичному періоді інтенсивно утворюються РНК і білок, а кількість білків-гістонів строго подвоюється.
Приблизно 1% ДНК тваринної клітини знаходиться в мітохондріях. Незначна частина мітохондріальної ДНК редупліціруется в синтетичному, тоді як основна - в постсинтетичному періоді інтерфази. Разом з тим відомо, що тривалість життя мітохондрій печінкових клітин, наприклад, становить 10 діб. З огляду на, що в звичайних умовах гепатоцити діляться рідко, слід допустити, що редуплікація ДНК мітохондрій може відбуватися незалежно від стадій мітотичного циклу.
Відрізок часу від закінчення синтетичного періоду до початку мітозу займає постсинтетичний (предмітотічноі), або G2-nepuod інтерфази. Він характеризується інтенсивним синтезом РНК і особливо білка. Завершується подвоєння маси цитоплазми в порівнянні з початком інтерфази. Це необхідно для вступу клітини в мітоз. Частина утворених білків (тубуліну) використовується в подальшому для побудови мікротрубочок веретена поділу. Синтетичний і постсинтетичний періоди пов'язані з митозом безпосередньо. Це дозволяє виділити їх в особливий період інтерфази - Препрофаза.
У мітозі можна виділити чотири фази. Головні події по окремих фаз представлені на рис. 2.13.
Поряд з перетворенням будови і впорядкованими переміщеннями хромосом обов'язковим для розділової фази циклу є побудова мітотичного апарату. Він складається з системи мікротрубочок (ахроматинового веретено, або веретено поділу світлооптичної мікроскопії) і структур, що поляризують мітоз1, т. е. що позначають два полюси в клітці, до яких розійдуться дочірні хромосоми. У клітинах ссавців і людини роль поляризують структур виконують центріолі.
Мітотичний апарат забезпечує спрямоване переміщення дочірніх хромосом в анафазе. Для цього необхідний контакт микротрубочек зі спеціалізованими ділянками
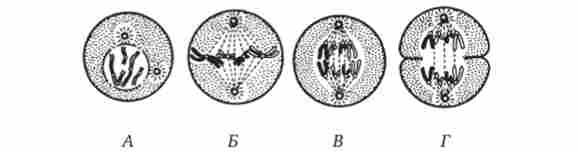
Мал. 2.13. Мітоз в тваринній клітині.
А - профази; Б - метафаза; В - анафаза; Г - телофаза
фаза мітозу
зміст змін
профаза
метафаза
анафаза
телофаза
Хромосоми спирализуются і набувають вигляду ниток. Ядро руйнується. Розпадається ядерна оболонка. У цитоплазмі зменшується кількість структур шорсткою мережі. Різко скорочується число полісом. Центриоли клітинного центру розходяться до полюсів клітини, між ними мікротрубочки утворюють веретено поділу
Закінчується утворення веретена поділу. Хромосоми шикуються в екваторіальній площині клітини (метафазної пластинка). Микротрубочки веретена поділу пов'язані з кінетохор хромосом. Кожна хромосома поздовжньо розщеплюється на дві хроматиди (дочірні хромосоми), з'єднані в області кінетохора
Зв'язок між хроматидами порушується, і вони в якості самостійних хромосом переміщаються до полюсів клітини зі швидкістю 0,2-5 мкм / хв. По завершенні руху на полюсах збирається два рівноцінних повних набору хромосом
Реконструюються інтерфазних ядра дочірніх клітин. Хромосоми деспирализуются. Утворюються ядерця. Руйнується веретено поділу. Материнська клітина ділиться на дві дочірні
хромосом - центромерами (кінетохор). При руйнуванні центромер в експерименті розбіжності хромосом до полюсів клітини не відбувається.
Порушення тієї чи іншої фази мітозу призводять до патологічних змін клітин. Відхилення від нормального перебігу процесу спирализации може привести до набухання і злипання хромосом. Іноді спостерігається відрив ділянки хромосоми, який, якщо він позбавлений центромери, не бере участі в анафазного переміщенні до полюсів і втрачається. Відставати при русі можуть окремі хроматиди, що призводить до утворення дочірніх ядер з незбалансованими хромосомними наборами. Пошкодження з боку веретена поділу призводять до затримки мітозу в метафазі, розсіюванню хромосом. При зміні кількості центриолей виникають багатополюсні або асиметричні мітози. Порушення цитотомії призводить до появи дво- і багатоядерних клітин.
Дані генетики і цитології вказують на збереження структурної індивідуальності хромосом у клітинному циклі. Є свідчення на користь упорядкованого розміщення хромосом в обсязі інтерфазних ядра. Особливості взаиморасположения хромосом можуть мати велике функціональне значення. Так, просторова близькість в клітинах людини хромосом 13, 14, 15, 21 і 22-й пар, що містять гени рРНК, пояснюється, по-видимому, їх участю у формуванні ядерця і освіті рибосом, а 11-й і 16-й хромосом - в освіті молекули гемоглобіну. Є вказівки на те, що в клітинах різних типів розташування хромосом не однаково.
На основі мітотичного циклу виник ряд механізмів, за допомогою яких в тому чи іншому органі кількість генетичного матеріалу і, отже, інтенсивність обміну можуть бути збільшені при збереженні сталості числа клітин. Подвоєння ДНК клітини не завжди супроводжується її поділом на дві. Оскільки механізм такого подвоєння збігається з предмітотічної редуплікацією ДНК і воно супроводжується кратним збільшенням кількості хромосом, це явище отримало назву ендомітозу.
З генетичної точки зору, Ендомітоз - геномна соматична мутація, про що буде сказано нижче. Інше явище, подібне по результату і назване політенії, полягає в кратному збільшенні вмісту ДНК в хромосомах при збереженні диплоидного кількості. Ендомітоз і політенія призводять до утворення поліплоїдних клітин, відрізняються кратним збільшенням обсягу спадкового матеріалу. У таких клітинах на відміну від диплоїдних гени повторені більш ніж два рази. Паралельно зі збільшенням числа генів зростає маса клітини, що підвищує її функціональні можливості. В організмі ссавців полиплоидизация з віком зустрічається в печінкових клітинах.