| Головна |
| «« | ЗМІСТ | »» |
|---|
Незважаючи на еволюційно відпрацьований механізм, що дозволяє зберігати постійної фізико-хімічну і морфологічну організацію хромосом у ряді клітинних поколінь, під впливом різних впливів ця організація може змінюватися. В основі зміни структури хромосоми, як правило, лежить первинне порушення її цілісності - розриви, які супроводжуються різними перебудовами, званими хромосомними мутаціями або аберацією.
Розриви хромосом відбуваються закономірно в ході кросинговеру, коли вони супроводжуються обміном відповідними ділянками між гомологами (див. Розд. 3.6.2.3). Порушення кросинговеру, при якому хромосоми обмінюються нерівноцінних генетичним матеріалом, призводить до появи нових груп зчеплення, де окремі ділянки випадають - делеции - або подвоюються - дуплікації (Рис. 3.57). При таких перебудовах змінюється число генів у групі зчеплення.
Розриви хромосом можуть виникати також під впливом різних мутагенних чинників, головним чином фізичних (іонізуючого та інших видів випромінювання), деяких хімічних сполук, вірусів.
Порушення цілісності хромосоми може супроводжуватися поворотом її ділянки, що знаходиться між двома розривами, на 180 ° - інверсія. Залежно від того, чи включає дану ділянку область центромери чи ні, розрізняють періцентрічні і парацентричної інверсії (див. рис. 3.57).
Фрагмент хромосоми, що відокремився від неї при розриві, може бути втрачений кліткою при черговому митозе, якщо він не має центромери. Найчастіше такий фрагмент прикріплюється до однієї з хромосом - транслокация. Нерідко дві пошкоджені негомологічної хромосоми взаємно обмінюються відірвалися ділянками - реципрокная транслокация (Див. Рис. 3.57). Можливо приєднання фрагмента до своєї ж хромосомі, але в новому місці - транспозиція (Див. Рис. 3.57). Таким чином, різні види інверсій і транслокаций характеризуються зміною локалізації генів.
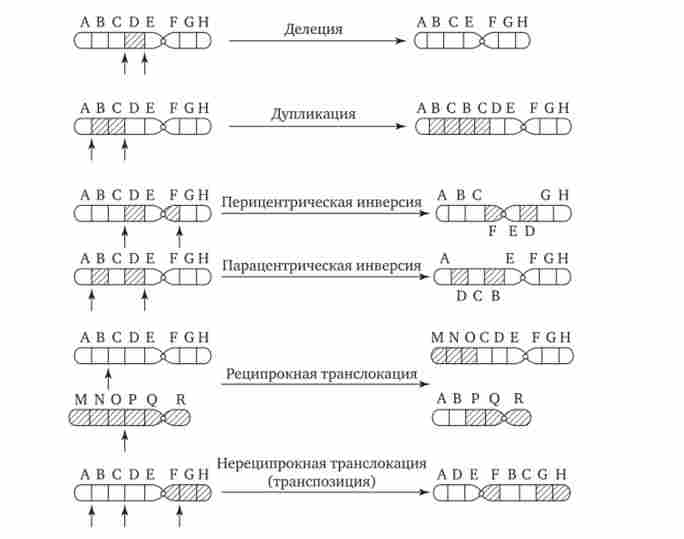
Мал. 3.57. Види хромосомних перебудов
Хромосомні перебудови, як правило, проявляються в зміні морфології хромосом, що можна спостерігати у світловий мікроскоп. Метацентрічна хромосоми перетворюються в субметацен- тричних і акроцентрічні, і, навпаки (рис. 3.58), з'являються кільцеві і полицентричні хромосоми (рис. 3.59). Особливу категорію хромосомних мутацій представляють аберації, пов'язані з центричним злиттям або поділом хромосом, коли дві негомологічних структури об'єднуються в одну-робертсонівські транслокация, або одна хромосома утворює дві самостійні хромосоми (рис. 3.60). При таких мутаціях не тільки з'являються хромосоми з новою морфологією, а й змінюється їх кількість в каріотипі.
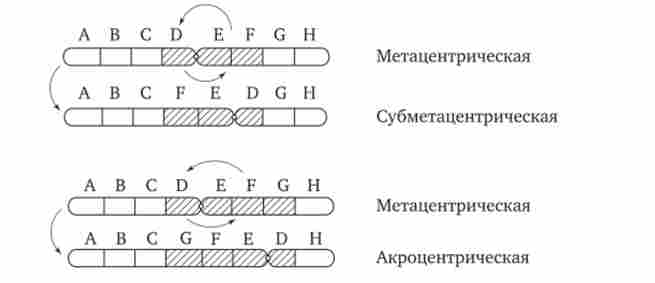
Мал. 3.58. Зміна форми хромосом в результаті періцентрічних інверсій
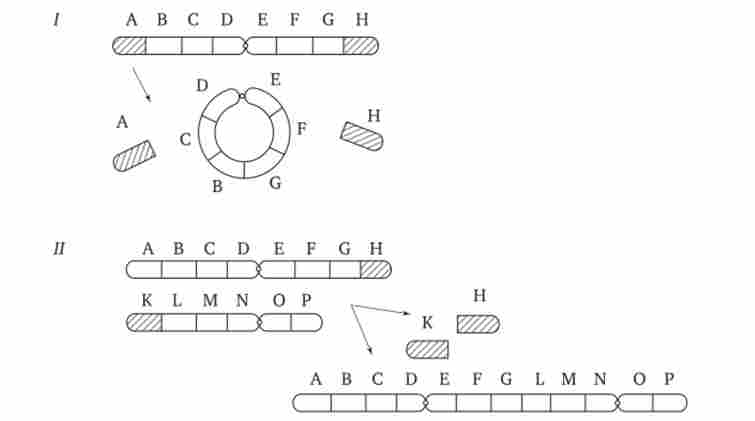
Мал. 3.59. Освіта кільцевих (/) і поліцентричної (//) хромосом
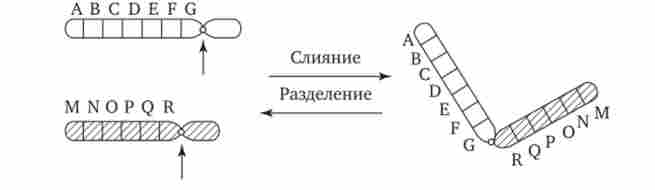
Мал. 3.60. Хромосомні перебудови, пов'язані з центричним злиттям або поділом хромосом, є причиною зміни числа хромосом в каріотипі
Описані структурні зміни хромосом, як правило, супроводжуються зміною генетичної програми, одержуваної клітинами нового покоління після поділу материнської клітини, так як змінюється кількісне співвідношення генів (при делециях і дуплікації), змінюється характер їх функціонування в зв'язку зі зміною взаємного розташування в хромосомі (при інверсії і транспозиції) або з переходом в іншу групу зчеплення (при транслокації). Найчастіше такі структурні зміни хромосом негативно позначаються на життєздатності окремих соматичних клітин організму, але особливо серйозні наслідки мають хромосомні перебудови, що відбуваються в попередниках гамет.
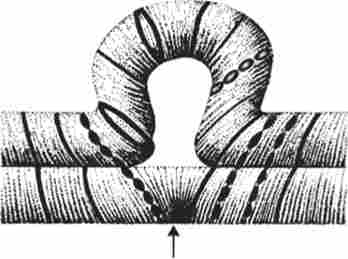
Мал. 3.61. Петля, що утворюється при кон'югації гомологічних хромосом, які несуть нерівноцінний спадковий матеріал у відповідних ділянках в результаті хромосомної перебудови
Зміни структури хромосом в попередниках гамет супроводжуються порушенням процесу кон'югації гомологів в мейозі і їх подальшого розбіжності. Так, делеции або дуплікації ділянки однієї з хромосом супроводжуються при кон'югації освітою петлі гомологом, що мають надлишкову матеріал (рис. 3.61). Реципрокная транслокация між двома негомологічної хромосоми призводить до утворення при кон'югації НЕ бивалента, а квадрівалента, в якому хромосоми утворюють фігуру хреста завдяки взаємному тяжінню гомологічних ділянок, розташованих в різних хромосомах (рис. 3.62). Участь в реципрокних транслокаціях більшого числа хромосом з утворенням полівалента супроводжується формуванням ще більш складних структур при кон'югації (рис. 3.63).
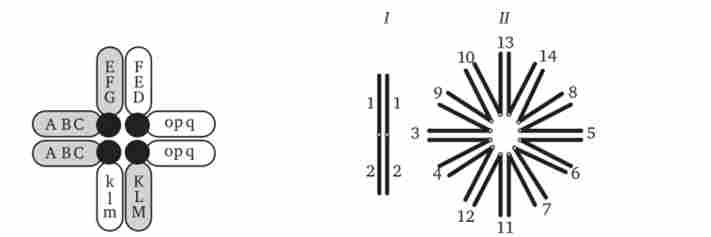
Мал. 3.62. Схема реципрокною транслокации між двома негомологічної хромосоми
Мал. 3.63. Освіта за кон'югації полівалента шістьма парами хромосом, що беруть участь в реципрокних транслокаціях.
/ - кон'югація між парою хромосом, що не несуть тран- слокацію; II - полівалент, утворений шістьма парами хромосом, що беруть участь в транслокації
У разі інверсії бивалент, що виникає в профазі I мейозу, утворює петлю, що включає взаємно інвертований ділянка (рис. 3.64).
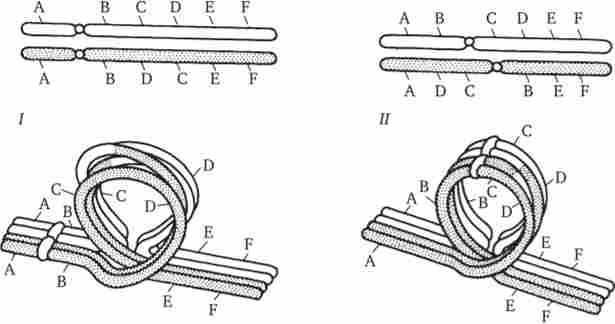
Мал. 3.64. Кон'югація хромосом при інверсіях.
I - парацентричної інверсія в одному з гомологів; II- періцентрічна інверсія в одному з гомологів
Кон'югація і подальше розходження структур, утворених зміненими хромосомами, призводить до появи нових хромосомних перебудов. В результаті гамети, отримуючи незбалансований спадковий матеріал, не здатні забезпечити формування нормального організму нового покоління. Причиною цього є порушення співвідношення генів, що входять до складу окремих хромосом, і їх взаємного розташування.
Однак, незважаючи на несприятливі, як правило, наслідки хромосомних мутацій, іноді вони виявляються сумісними з життям клітини і організму і забезпечують можливість еволюції структури хромосом, що лежить в основі біологічної еволюції. Так, невеликі за розміром делеції можуть зберігатися в гетерозиготному стані в ряду поколінь. Менш шкідливими, ніж делеции, є дуплікації, хоча великий обсяг матеріалу в збільшеній дозі (більше 10% генома) призводить до загибелі організму.
Нерідко життєздатними виявляються робертсонівські транслокации, часто не зв'язані зі зміною обсягу спадкового матеріалу. Цим можна пояснити варіювання числа хромосом в клітинах організмів близькоспоріднених видів. Наприклад, у різних видів дрозофіли кількість хромосом в гаплоидном наборі коливається від 3 до 6, що пояснюється процесами злиття і поділу хромосом. Можливо, істотним моментом у появі виду Homo sapiens були структурні зміни хромосом у його мавпоподібних предків. Встановлено, що два плеча великої другий хромосоми людини відповідають двом різним хромосомами сучасних людиноподібних мавп (12-й і 13-й - шимпанзе, 13-й і 14-й - горили і орангутана). Ймовірно, ця людська хромосома утворилася в результаті центричного злиття за типом робертсоновской транслокации двох мавпячих хромосом.
До істотного варіювання морфології хромосом, який лежить в основі їх еволюції, призводять транслокации, транспозиції і інверсії. Аналіз хромосом людини показав, що його 4, 5,12 і 17-я хромосоми відрізняються від відповідних хромосом шимпанзе періцентрічними інверсіями.
Таким чином, зміни хромосомної організації, найчастіше що погано впливають на життєздатність клітини і організму, з певною ймовірністю можуть бути перспективними, успадковуватися в ряду поколінь клітин і організмів і створювати передумови для еволюції хромосомної організації спадкового матеріалу.