| Головна |
| «« | ЗМІСТ | »» |
|---|
Нескоректована зміни хімічної структури генів, що відтворюються в послідовних циклах реплікації і проявляються у потомства у вигляді нових варіантів ознак, називають генними мутаціями.
Зміни структури ДНК, що утворює ген, можна розділити на три групи. Мутації першої групи полягають в заміні одних підстав іншими. Вони складають близько 20% спонтанних генних змін. Друга група мутацій обумовлена зсувом рамки зчитування, що відбувається при зміні кількості нуклеотидних пар в складі гена. Нарешті, третю групу представляють мутації, пов'язані зі зміною порядку нуклеотиднихпослідовностей в межах гена (інверсії).
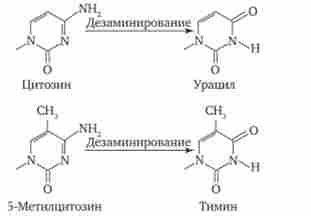
Мал. 3.18. Спонтанне дезамінування цитозину
Мутації за типом заміни азотистих основ відбуваються в силу ряду конкретних причин. Однією з них може бути виникає випадково або під впливом конкретних хімічних агентів зміна структури підстави, вже включеного в спіраль ДНК. Якщо така змінена форма підстави залишається непоміченою ферментами репарації, то при найближчому циклі реплікації вона може приєднувати до себе інший нуклеотид. Прикладом може служити дезаминирование цитозину, що перетворюється в урацил мимовільно або під впливом азотної кислоти (рис. 3.18). Утворений при цьому урацил, не помічений ферментом ДНК-глікозілазой, при реплікації з'єднується з аде- нином, який згодом приєднує тіміділових нуклеотид. В результаті пара Ц-Г заміщається в ДНК парою Т-А (рис. 3.19, Г). Дезаминирование метилованого цитозину перетворює його в тимін (див. Рис. 3.18). Тіміділових нуклеотид, будучи природним компонентом ДНК, не виявляється ферментами репарації як зміна і при наступній реплікації приєднує аде- Нілов нуклеотид. В результаті замість пари Ц-Г в молекулі ДНК також з'являється пара Т-А (рис. 3.19, II).
Іншою причиною заміни підстав може бути помилкове включення в синтезируемую ланцюг ДНК нуклеотиду, що несе хімічно змінену форму підстави або його аналог. Якщо ця помилка залишається непоміченою ферментами реплікації і репарації, змінене підставу включається в процес реплікації, що нерідко призводить до заміни однієї пари на іншу. Прикладом цього може служити приєднання в ході реплікації до аденіну материнської ланцюга нуклеотиду з 5-бромурацілом (5-БО), аналогічного тіміділових нуклеотиду. При подальшій реплікації 5-БО приєднує до себе не аденін, а гуанін. Гуанін в ході подальшого подвоєння утворює комплементарних пару з цитозином. В результаті пара А-Т замінюється в молекулі ДНК парою Г-Ц (рис. 3.20).
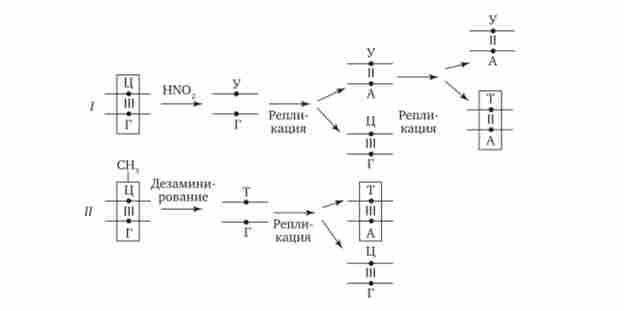
Мал. 3.19. Мутації за типом заміни підстави (дезаминирование азотистих основ в ланцюзі ДНК).
I - перетворення цитозину в урацил, заміна Ц-Г-пари на Т-А-пару; II - перетворення метілцітозіна в тимін, заміна Ц-Г-пари на Т-А-пару
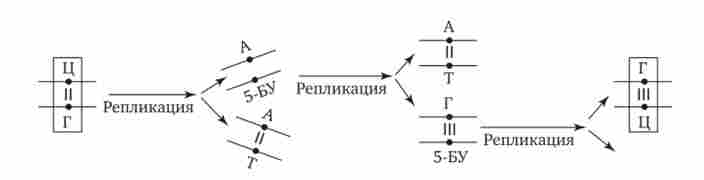
Мал. 3.20. Мутації за типом заміни підстав (включення аналога азотистого підстави при реплікації ДНК)
З наведених прикладів видно, що зміни структури молекули ДНК за типом заміни підстав виникають або до, або під час реплікації спочатку в одній полінуклеотидних ланцюга. Якщо такі зміни не виправляються в ході репарації, то при подальшій реплікації вони стають надбанням обох ланцюгів ДНК.
Наслідком заміни однієї пари комплементарних нуклеотидів на іншу є утворення нового триплетів в нуклеотидної послідовності ДНК, що кодує послідовність амінокислот у пептидного ланцюга. Це може і не відбитися на структурі пептиду в тому випадку, якщо новий триплет буде «синонімом» колишнього, т. Е. Буде кодувати ту ж амінокислоту. Наприклад, амінокислота валін шифрується чотирма триплету: ЦАА, ЦАГ, ЦАТ, цяць. Заміна третього підстави в будь-якому з цих триплетів не змінить його сенсу (вирожденність генетичного коду).
У тому випадку, коли знову виник триплет шифрує іншу амінокислоту, змінюються структура пептидного ланцюга і властивості відповідного білка. Залежно від характеру і місця трапилася заміни специфічні властивості білка змінюються в різному ступені. Відомі випадки, коли заміна лише однієї амінокислоти в пептиді істотно впливає на властивості білка, що проявляється в зміні більш складних ознак. Прикладом може служити зміна властивостей гемоглобіну людини при серповидно-клітинної анемії (рис. 3.21). В такому гемоглобіні - (HbS) на відміну
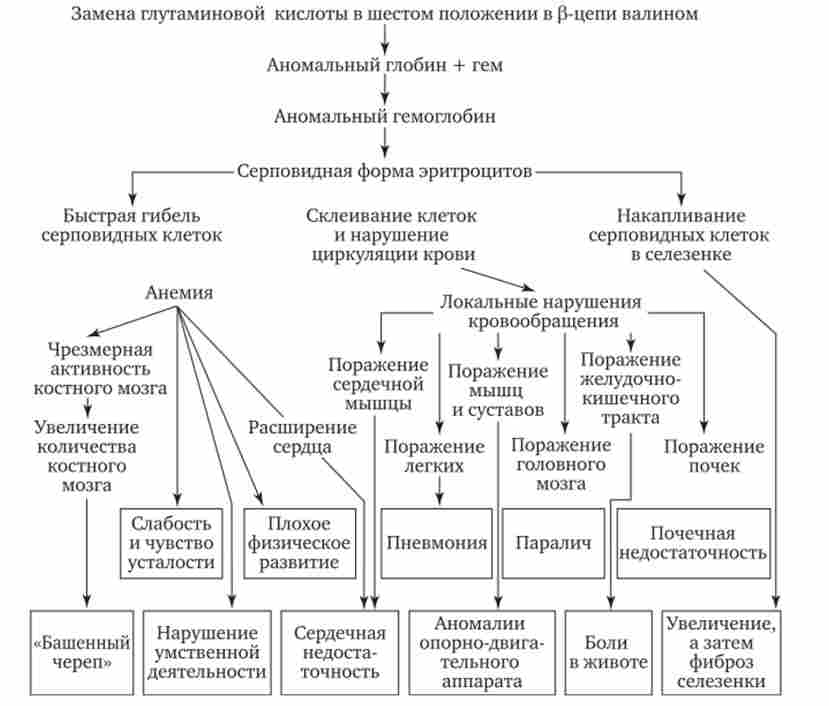
Мал. 3.21. Пдейотропний ефект заміни однієї амінокислоти в (3-ланцюга гемоглобіну людини, що призводить до розвитку серповидно-клітинної анемії від нормального НЬА - в (3-глобінових ланцюгах в шостому положенні глутамінова кислота замінена валіном. Це є наслідком заміни одного з підстав в триплеті, шифр глутаминовую кислоту (ЦТТ або ЦТЦ). в результаті з'являється триплет, шифрувальний валін (ЦАТ або цяць). в даному випадку заміна однієї амінокислоти в пептиді істотно змінює властивості глобина, що входить до складу гемоглобіну (знижується його спосіб ність зв'язуватися з 02), У людини розвиваються ознаки серповидно-клітинної анемії.
У деяких випадках заміна одного підстави на інше може привести до появи одного з нонсенс-триплетів (ATT, АТЦ, АЦТ), що не шифрувального ніякої амінокислоти. Наслідком такої заміни буде переривання синтезу пептидного ланцюга. Підраховано, що заміни нуклеотидів в одному триплети призводять в 25% випадків до утворення триплетів-синонімів; в 2-3 - безглуздих триплетів, в 70-75% - до виникнення справжніх генних мутацій.
Таким чином, мутації за типом заміни підстав можуть виникати як в результаті спонтанних змін структури підстави в одному з ланцюжків вже існуючої подвійної спіралі ДНК, так і в ході реплікації у знову синтезованої ланцюга. У тому випадку, якщо ці зміни не виправляються в процесі репарації (або, навпаки, виникають в ході репарації), вони фіксуються в обох ланцюгах і далі будуть відтворюватися в наступних циклах реплікації. Отже, важливим джерелом виникнення таких мутацій є порушення процесів реплікації і репарації.
Мутації зі зсувом рамки зчитування составлют значну частку спонтанних мутацій. Вони відбуваються внаслідок випадання або вставки в нуклеотидну послідовність ДНК однієї або декількох пар комплементарних нуклеотидів. Велика частина вивчених мутацій, що викликають зрушення рамки, виявлена в послідовності, що складаються з однакових нуклеотидів.
Зміні числа нуклеотидних пар в ланцюзі ДНК сприяють впливу на генетичний матеріал деяких хімічних речовин, наприклад акридиновим з'єднань. Деформуючи структуру подвійної спіралі ДНК, вони призводять до вставки додаткових підстав або їх випадання при реплікації. Прикладом служать мутації, отримані у фага Т4 при впливі профлавіна. Вони складаються у включенні або видаленні всього однієї нуклеотидної пари. Важливою причиною зміни кількості нуклеотидних пар в гені за типом великих делеций (випадінь) може бути рентгенівське опромінення. У плодової мухи, наприклад, відома мутація гена, контролюючого забарвлення очей, яка викликається опроміненням і складається в делеции близько 100 нуклеотидних пар.
Велике число мутацій за типом вставок відбувається внаслідок включення в послідовність нуклеотидів рухомих генетичних елементів - транспозони. транспозони - це досить протяжні нуклеотидні послідовності, вбудовані в геноми ЕУ і прокаріотів клітин, здатні спонтанно змінювати своє положення (див. розд. 3.6.4.3).
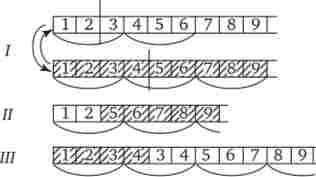
Мал. 3.22. Мутації зі зсувом рамки зчитування (нерівноцінний обмін при вну трігенном кроссинговере).
З певною вірогідністю вставки і делеції можуть виникати в результаті помилок рекомбінації при нерівноцінних вну трігенном кроссинговере (рис. 3.22).
I-розриви алельних генів в різних ділянках і обмін фрагментами між ними; II - випадання 3-й і 4-й пар нуклеотидів, зрушення рамки зчитування; III - подвоєння 3-й і 4-й пар нуклеотидів, зрушення рамки зчитування
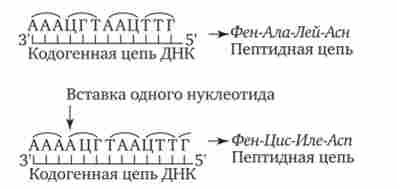
Мал. 3.23. Слідство зміни кількості нуклеотидних пар в молекулі ДНК.
Зрушення рамки зчитування в результаті вставки одного нуклеотиду в кодогенную ланцюг призводить до зміни складу зашифрованого в ній пептиду
При безперервності зчитування і неперекриваемості генетичного коду зміна кількості нуклеотидів, як правило, призводить до зсуву рамки зчитування і зміни змісту біологічної інформації, записаної в даній послідовності ДНК (рис. 3.23). Однак, якщо кількість вставлених або втрачених нуклеотидів кратно трьом, зсуву рамки може не відбутися, але це призведе до включення додаткових амінокислот або випадання частини їх з поліпеп- тідной ланцюга. Можливим наслідком зсуву рамки є виникнення нонсенс-триплетів, що веде до синтезу укорочених пептидних ланцюгів.
Мутації за типом інверсії нуклеотидних послідовностей в гені. Даний тип мутацій відбувається внаслідок повороту ділянки ДНК на 180 °. Зазвичай цьому передує освіту молекулою ДНК петлі, в межах якої реплікація йде в напрямку, протилежному правильному.
В межах інвертованого ділянки порушується зчитування інформації, в результаті змінюється і амінокислотна послідовність білка.