| Головна |
| «« | ЗМІСТ | »» |
|---|
Для підтримки основних параметрів клітини або організму протягом їх життя, а також в ряді поколінь спадковий матеріал повинен відрізнятися стійкістю до зовнішніх впливів або повинні існувати механізми корекції виникаючих в ньому змін. У живій природі використовуються обидва чинники. Третім фактором є точність копіювання нуклеотиднихпослідовностей материнської ДНК в процесі її реплікації.
За реакційної здатності молекули ДНК відносяться до категорії хімічно інертних речовин. Відомо, що роль речовини спадковості може виконувати не тільки ДНК, але і РНК (деякі віруси). Вважають, що вибір на користь ДНК обумовлений її нижчою в порівнянні з РНК реакційною здатністю.
Розглянутий вище механізм реплікації відрізняється надзвичайно високою точністю відтворення структури ДНК. При подвоєнні ДНК помилки виникають в середньому з частотою 1 - 10 ~6 комплементарних пар основ.
У підтримці високої точності реплікації важлива роль належить насамперед ферменту ДНК-полімерази. Цей фермент здійснює відбір необхідних нуклеотидів з числа наявних у ядерному соку нуклеозидтрифосфатів (АТФ, ТТФ, ГТФ, ЦТФ), точне приєднання їх до матричного ланцюга ДНК і включення в зростаючу дочірню ланцюг (див. Рис. 3.10). Частота включення неправильних нуклеотидів на цій стадії становить 1 - 10 "5 пар основ.
Такі помилки в роботі ДНК-полімерази пов'язані з виникненням змінених форм азотистих основ, які утворюють «незаконні» пари з підставами материнської ланцюга. Наприклад, змінена форма цитозину замість гуаніну зв'язується водневими зв'язками з аденін. В результаті в зростаючу ланцюг ДНК включається помилковий нуклеотид. Швидкий перехід зміненої форми такої підстави в звичайну порушує його зв'язування з матрицею, з'являється неспарених З'-ОН-кінець ланцюга ДНК. У цій ситуації включається механізм самокорекції, здійснюваний ДНК-полімеразою (або тісно пов'язаних з нею ферментом - редагую ендонуклеази). Самокорекція полягає в відщепленні помилково включеного в ланцюг ДНК нуклеотиду, що не спареного з матрицею (рис. 3.14). Наслідком самокорекції є зниження частоти помилок в 10 разів (з 10 ~5 до 10 ~6).
Незважаючи на ефективність самокорекції, в ході реплікації після подвоєння ДНК в ній виявляються помилки. Особливо часто це спостерігається при порушенні концентрації чотирьох нуклеозидтрифосфатів в навколишньому субстраті. Значна частина змін виникає також в молекулах ДНК в результаті спонтанно відбуваються, пов'язаних з втратою пуринових підстав - аденіну і гуаніну (апурінізаціей) - або дезаминированием цитозину, який перетворюється в урацил. Частота останніх змін досягає 100 на 1 геном / сут.
Вміщені в ДНК підстави можуть змінюватися під впливом реакційноздатних сполук, які порушують їх нормальне спаровування, а також під дією ультрафіолетового випромінювання, яке може викликати утворення ковалентного зв'язку між двома сусідніми залишками тиміну в ДНК (димери тиміну). Названі зміни в черговому циклі реплікації повинні привести або до випадання пар основ в дочірньої ДНК, або до заміни одних пар іншими. Зазначені зміни дійсно супроводжують
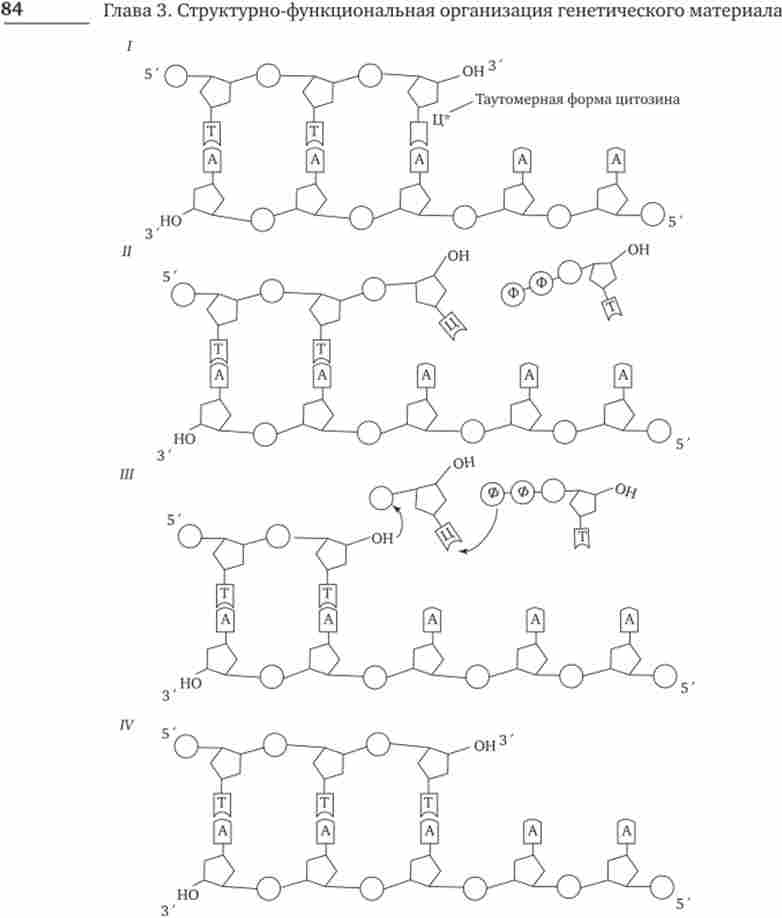
Мал. 3.14. Схема процесу корекції при синтезі ДНК.
I - включення в ланцюг ДНК нуклеотиду зі зміненою (таутомерну) формою цитозину, який «незаконно» злучається з аденін; II - швидкий перехід цитозину в звичайну форму порушує його спаровування з аденін; неспарених З'-ОН-кінець синтезується ланцюга перешкоджає подальшому її подовження під дією ДНК-полімерази; III - ДНК-полімераза видаляє незаконний нуклеотид, в результаті чого знову з'являється спарений з матрицею З'-ОН-кінець; / V - ДНК полімераза продовжує нарощування ланцюга на З'-ОН-кінці кожен цикл реплікації ДНК, проте їх частота значно менше, ніж повинна була б бути. Це пояснюється тим, що більшість змін такого роду усувається завдяки дії механізму репарації (Молекулярного відновлення) вихідної нуклеотидної послідовності ДНК.
Механізм репарації заснований на наявності в молекулі ДНК двох комплементарних ланцюгів. Спотворення послідовності нуклеотидів в одній з них виявляється специфічними ферментами. Потім відповідну ділянку видаляється і заміщується новим, синтезованим на другий комплементарного ланцюжка ДНК. Таку репарацію називаютьексцизійної, т. е. з «вирізанням» (рис. 3.15). Вона здійснюється до чергового циклу реплікації, тому її називають також дореплікатівной.
Відновлення вихідної структури ДНК вимагає участі ряду ферментів. Важливим моментом в запуску механізму репарації є виявлення помилки в структурі ДНК. Нерідко такі помилки виникають у знову синтезованої ланцюга в процесі реплікації. Ферменти репарації повинні виявити саме цю ланцюг. У багатьох видів живих організмів знову синтезована ланцюг ДНК відрізняється від материнської ступенем метилювання її азотистих основ, яке відстає від синтезу. Репарації при цьому піддається неметілірованная ланцюг. Об'єктом впізнавання ферментами репарації можуть також служити розриви в ланцюзі ДНК. У вищих організмів, де синтез ДНК відбувається безупинно, а окремими реплі- конамі, знову синтезируемая ланцюг ДНК має розриви, що робить можливим її впізнавання.
Відновлення структури ДНК при втраті пуринових підстав однієї з її ланцюгів передбачає виявлення дефекту за допомогою ферменту ендонуклеази, яка розриває фосфоефірную зв'язок
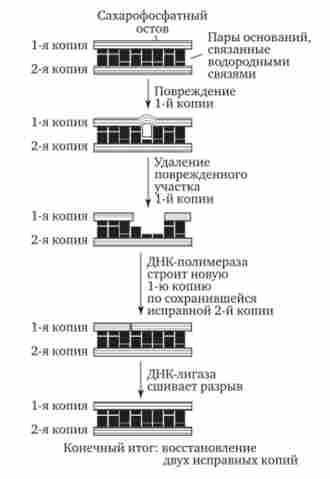
Мал. 3.15. Схема ексцизійної, дореплікатівной репарації ДНК
в місці пошкодження ланцюга. Потім змінений ділянку з декількома примикають до нього нуклеотидами видаляється ферментом екзонуклеаза, а на його місці відповідно до порядку підстав комплементарного ланцюжка утворюється правильна нуклеотидних послідовність (рис. 3.15).
При зміні одного з підстав в ланцюзі ДНК у відновленні вихідної структури беруть участь ферменти ДНК-глікозілази числом близько 20. Вони специфічно впізнають пошкодження, обумовлені дезаминированием, алкилированием і іншими структурними перетвореннями підстав. Такі модифіковані підстави видаляються. Виникають ділянки, позбавлені підстав, які репаруючу, як при втраті пуринів. Якщо відновлення нормальної структури не здійснюється, наприклад в разі дезаминирования азотистих основ, відбувається заміна одних пар комплементарних основ іншими - пара Ц-Г може замінюватися парою Т-Аіт. п. (див. розд. 3.4.2.3).
Освіта в полінуклеотидних ланцюгах під дією УФ-променів тимінових димарів (Т-Т) вимагає участі ферментів, котрі дізнаються не окремі змінені підстави, а більш протяжні пошкодження структури ДНК. Репаративний процес в цьому випадку також пов'язаний з видаленням ділянки, що несе димер, і відновленням нормальної послідовності нуклеотидів шляхом синтезу на комплементарного ланцюжка ДНК.
У тому випадку, коли система ексцизійної репарації не виправляє зміни, що виник в одного ланцюга ДНК, в ході реплікації відбувається фіксація цієї зміни і воно стає надбанням обох ланцюгів ДНК. Це призводить до заміни однієї пари комплементарних нуклеотидів на іншу або до появи розривів (проломів) у знову синтезованої ланцюга проти змінених ділянок. Відновлення нормальної структури ДНК при цьому може статися і після реплікації.
постреплікатівной репарація здійснюється шляхом рекомбінації (обміну фрагментами) між двома новоствореними подвійними спіралями ДНК. Прикладом такої постреплікатівной репарації може служити відновлення нормальної структури ДНК при виникненні тимінових димарів (Т-Т), коли вони не усуваються мимовільно під дією видимого світла (Світлова репарація) Або в ході дореплікатівной ексцизійної репарації.
Ковалентні зв'язки, що виникають між рядом стоять залишками тиміну, роблять їх нездатними до зв'язування з комплементарними нуклеотидами. В результаті у знову синтезованої ланцюга ДНК з'являються розриви (проломи), впізнавані ферментами репарації. Відновлення цілісності нової полінуклеотидних ланцюга однієї з дочірніх ДНК здійснюється завдяки рекомбінації з відповідною їй нормальної материнської ланцюгом іншої дочірньої ДНК. Утворився в материнській ланцюга пробіл заповнюється потім шляхом синтезу на комплементарної їй полінуклеотидних ланцюга (рис. 3.16). Проявом такої постреплікатівной репарації, здійснюваної шляхом рекомбінації між ланцюгами двох дочірніх молекул ДНК, можна вважати нерідко спостерігається обмін матеріалом між сестринськими хроматидами (рис. 3.17).
В ході дореплікатівной і постреплікатівной репарації відновлюється велика частина пошкоджень структури ДНК. Однак, якщо в спадковому матеріалі клітини виникає занадто багато пошкоджень і частина з них не ліквідується, включається система індукованих (спонукує) ферментів репарації (SOS-система). Ці ферменти заповнюють прогалини, відновлюючи цілісність синтезованих полінуклеотидних ланцюгів без точного дотримання принципу комплементарності. Ось чому іноді самі процеси репарації можуть служити джерелом стійких змін в структурі ДНК (мутацій). Названа реакція також відноситься до SOS-системі.
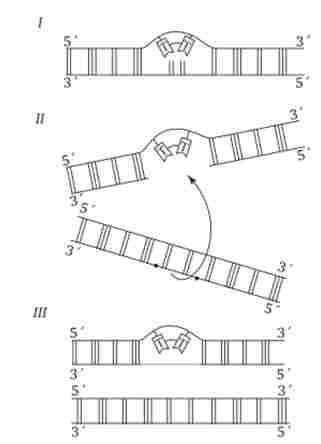
Мал. 3.16. Схема постреплікатівной репарації ДНК.
I-виникнення тимінових димера в одному з ланцюжків ДНК;
II - утворення «дірки» у знову синтезованої ланцюга проти зміненого ділянки материнської молекули після реплікації (стрілкою показано подальше заповнення «дірки» ділянкою з відповідним ланцюгом другий дочірньої молекули ДНК);
III - відновлення цілісності дочірньої ланцюга верхній молекули за рахунок рекомбінації і в нижній молекулі за рахунок синтезу на комплементарного ланцюжка
Якщо в клітці, незважаючи на здійснювану репарацію, кількість пошкоджень структури ДНК залишається високим, в ній блокуються процеси реплікації ДНК. Така клітина не ділиться, а значить, не передає виникли змін потомству.
Її викликає ушкодженнями ДНК зупинка клітинного циклу в поєднанні з неможливістю молекулярної репарації зміненого спадкового матеріалу може за участю білка, синтез якого контролюється геном р53, приводити до активації процесу самоліквідації (апоптоз) дефектної клітини з метою усунення її з організму.
Таким чином, великий набір різних ферментів репарації здійснює безперервний «огляд» ДНК, видаляючи з неї пошкоджені ділянки і сприяючи підтримці стабільності спадкового матеріалу.
Спільна дія ферментів реплікації (ДНК-полімераза і редагує ендонуклеаза) і ферментів оепаіаііі забезпечує досить низьку частоту

Мал. 3.17. Межхроматідние обміни (вказані стрілками) помилок в молекулах ДНК, яка підтримується на рівні 1 - 10-9 пар змінених нуклеотидів на геном. При розмірі генома людини 3 - 109 нуклеотидних пар це означає появу близько 3 помилок на реплицирующихся геном. Разом з тим навіть цей рівень достатній для освіти за час існування життя на Землі значного генетичного різноманіття у вигляді генних мутацій.