| Головна |
| «« | ЗМІСТ | »» |
|---|
Синтез нуклеїнових кислот Дик і РНК з нуклеотидів відбувається в ядрі клітини і мітохондріях.
Синтез ДНК носить назву реплікація, т. е. створення «дочірніх» копій- реплік, ідентичних «батьківської» ДНК.
Синтез РНК носить назву транскрипція - «Переписування» інформації з матриці ДНК на матричну РНК (мРНК). Матрична РНК є вихідною молекулою для передачі інформації про структуру білків, які синтезуються в плазмі. Тому її називають також інформаційної РНК (іРНК).
В клітці є декілька видів РНК. Всі вони беруть участь в синтезі білка. Один з видів РНК носить назву транспортної (тРНК). Вона здійснює транспорт амінокислот з цитоплазми до рибосомальної РНК. Рибосомальна РНК (рРНК) входить до складу рибосом і контролює біосинтез білка. рРНК має більш варіабельний склад, ніж матрична або транспортна.
Практично всі живі організми, за винятком деяких видів бактерій, мають здатність синтезувати потрібні їм нуклеїнові кислоти.
Матричні, або інформаційні, РНК синтезуються на одній з ділянок молекули ДНК. Такі ділянки ДНК називають генами. Ген - це ділянка ДНК, що кодує інформацію про конкретний ознаці через послідовність нуклеотидів.
Роль матриць грають розділені ланцюга двунітевой материнської ДНК. Реплікація являє собою реакцію полімеризації мононуклеотидів Nub NU2, NU3 і т. Д. В дочірню підлогу і нуклеотидную ланцюг (див. Розд. 8.5) в послідовності, заданої генами:
Основну роль в реплікації ДНК відіграє фермент ДНК-полімераза, який з'єднує (полимеризует) мононуклеотиди в ланцюг дочірньої ДНК-репліки.
Полімеризаційного процес реплікації батьківських ДНК включає наступні стадії (рис. 9.27).
Для будь-якого синтезу органічних полімерів, здійснюваного in vitro або in vivo, потрібна енергія. На реакції полімеризації мононуклеотидів йде енергія, що звільняється всіма типами дезоксірібонуклсозідтріфосфатов, що беруть участь в синтезі ДНК. Утворений пирофосфат РР, під дією пірофосфатази також розщеплюється на дві молекули ортофосфата Р "даючи додаткову енергію для біосинтезу ДНК. Після реплікації дочірні спіралі закручуються назад вже без витрат енергії і будь-яких ферментів.
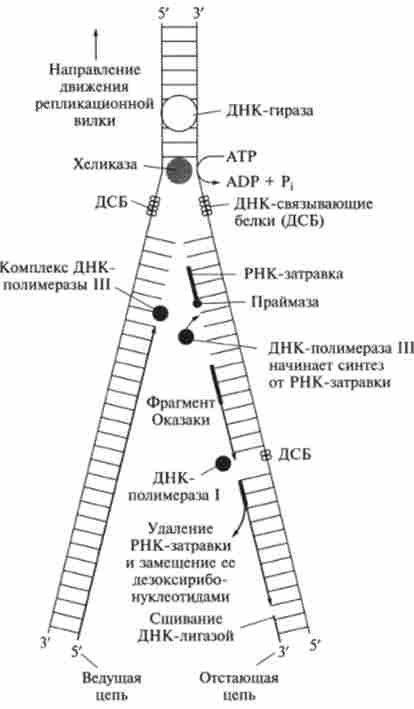
Мал. 9.27. Стадії реплікації ДНК (репликативная вилка)
Крім ДНК-полімерази в реплікації беруть участь більше 40 ферментів і білкових факторів, об'єднаних в загальну ДНК-репліказную систему, звану реплісомой. Ферменти ХЕЛІКАЗИ, топоізомераза і ДНК-зв'язуючі білки розплітає ДНК, утримують матрицю в розведеному стані і обертають молекулу ДНК.
Є кілька видів ДНК-полімерази, які каталізують реплікацію.
Першою була відкрита ДНК-полімераза I в кишкової палички Е. coli. Пізніше був відкритий мутант Е. coli, позбавлений ДНК-полімерази I, але здатний синтезувати ДНК за допомогою ДНК-полімерази II. Виявилося, що для реплікації ДНК Е. coli необхідна участь декількох ферментів. ДНК-полімераза I не наділено здатністю ініціювати синтез ланцюгів ДНК de novo.
Одним з добре вивчених ферментів, які беруть участь в стадії ініціації реплікації ДНК, є специфічна клітинна РНК-полімераза, названа праймазой. Праймаза каталізує синтез короткого олігорібонуклеотіда (від 10 до 60 нуклеотидів), т. Е. Праймера, з якого починається синтез ДНК. Іраймази розрізняються як за структурою, так і за специфічності дії.
Відкрито також комплекс білків dna В і dna С, який поблизу реплікаціон- ної вилки періодично бере участь у формуванні специфічної вторинної структури ДНК, що підходить для впізнавання праймазой.
Основним ферментом, що каталізує біосинтез новоствореної ДНК (точніше, стадію елонгації реплікації ДНК), є ДНК-полімераза III, яка представляє собою мультімерной комплекс ДНК-полімерази (молекулярна маса близько 900 000) і ряду інших білків. ДНК-полімераза III з Е. coli складається мінімум з 10 субодиниць.
Є докази, що в димерной формі ДНК-полімераза III каталізує пов'язаний синтез провідної (лідируючої) і відстає ланцюгів ДНК при реплікації. Більш точно з'ясовано також роль ДНК-полімерази I: вона каталізує відщеплення затравочного олігорібонуклеотідного праймера і заповнення що утворюються після цього прогалин (ніш) дезоксірібонуклеотідов. Відомо, що ДНК-полімераза II з Е. coli (Молекулярна маса 88 000) виконує «ремонтні» функції, виправляючи ушкодження ланцюгів ДНК.
Істотно, що ДНК-полімераза I в якості матриці використовує одноцепочечниє ділянки, в той час як ДНК-полімераза III - дволанцюжкові, в яких є короткі одноцепочечниє послідовності.
Важливу функцію з'єднання двох ланцюгів ДНК або замикання двох кінців одного ланцюга ДНК в процесі реплікації або репарації ДНК виконує особливий фермент - ДНК-лігаза, що каталізує за рахунок енергії АТФ освіту Фос фодіефірной зв'язку між ОН-групою дезоксирибози одного ланцюга і фосфатної групою інший ланцюга ДНК .
Функцію розкручування (розплітання) подвійної спіралі ДНК в реплікаціон- ної вилці виконує специфічний білок, названий ХЕЛІКАЗОЙ (молекулярна маса 300 000). Одноцепочечниє ділянки ДНК служать в якості матриці при реплікації. Вони стабілізуються за допомогою особливих білків (ДНК-зв'язуючі білки, молекулярна маса 75 600), що перешкоджають зворотному комплементарному взаємодії ланцюгів ДНК, в зв'язку з чим ці білки іноді називають дестабілізуючими подвійну спіраль білками.
Особливу роль в с верху пі рал і заці і грають ферменти топоізомерази, які забезпечують як реплікацію, так і транскрипцію ДНК. Ці ферменти наділені здатністю не тільки створювати супервитки, але і знищувати суперспіралі- зацию шляхом зшивання утворюються розривів або розрізання ДНК.
Нарешті, відкриті спеціальні ферменти, «редагують» ДНК. Вони здійснюють вирізання та видалення помилково включених нуклеотидів або усувають (репаруючу) пошкодження ДНК, викликані фізичними або хімічними факторами (рентгенівське випромінювання, УФ-промені, хімічний мутагенез).
До теперішнього часу в клітинах тварин, як і у бактерій, відкрито кілька ДНК-полімерази. У реплікації ДНК еукаріот беруть участь два основних типи полимераз - пі ГВ. Показано, що ДНК-полімераза п складається з 4 субодиниць і ідентична за структурою та властивостями у всіх клітинах ссавців, причому одна з субодиниць виявилася наділеною Праймазная активністю. Найбільша субодиниця ДНК-полімерази (молекулярна маса 180 000) каталізує реакцію полімеризації, в основному синтез відстає ланцюга ДНК, що є складовою частиною праймасоми. ДНК-полімераза ГВ складається з 2 субодиниць і переважно каталізує синтез провідної ланцюга ДНК (див. Далі). Відкрита також ДНК-полімераза ГС, яка в ряді випадків замінює ГВ-фермент, зокрема при репарації ДНК (виправлення порушень ДНК, викликаних помилками реплікації або повреждающими агентами).
Слід зазначити, що в клітині є два білкових фактора реплікації: RFA і RFC. Фактор реплікації RFA пов'язує одноцепочечную ДНК (на зразок білкових факторів зв'язування роз'єднаних ланцюгів ДНК при реплікації у Е. coli)y фактор RFC - стабілізатор всього Реплікаційний комплексу.
На першому етапі транскрипції ДНК-полімераза починає розплітати подвійну нитку ДНК і утворюється репликационная (репликативная) вилка (див. Рис. 9.27).
Синтез ДНК в клітинах людини і тварин відбувається в період клітинного циклу, званого S-фазою.
Реплікація ДНК починається відразу в декількох тисячах молекул в затравочних ділянках ДНК - праймерів, синтезованих спеціальними ферментами. Запал синтезується з рібонуклеозідфосфатов, відповідно ДНК закінчує синтез нового ланцюга, дійшовши до РНК-затравки. Наслідком цього є утворення фрагментів Окадзакі.
Щоб забезпечити спрямований синтез ланцюга ДНК, в дію всту пает особлива система репарацій, що видаляє РНК-затравки шляхом гідролізу і відновлює правильну струтури ДНК. Фрагмент завершує ДНК-лігаза, яка з'єднує кінець 3 'нового фрагмента з кінцем 5' попереднього.
В ході транскрипції подвійна нитка ДНК продовжує розплітатися і вилка переміщається від одного кінця молекули ДНК до іншого кінця нуклеотид за нуклеотидом: Nu, -> Nu, + j.
Ланцюги молекули ДНК розходяться, і кожна з них стає матрицею, на якій синтезується нова комплементарна ланцюг. В результаті утворюються нові двоспіральні молекули ДНК, ідентичні батьківській молекулі.
Нові двоспіральні молекули ДНК складаються з одного ланцюга початкової батьківської молекули і однієї знову синтезованою ланцюга.
Склад і послідовність розташування нуклеотидів в батьківської ДНК «переписуються» на дочірню ДНК за тим же принципом комплементарності, на основі якого утворюється двухспіральная молекула ДНК.
Згідно з принципом комплементарності вільні нуклеотиди А, Г, Т, Ц (в латинському написанні відповідно A, G, Т, С) приєднуються до нитки ДНК через водневі зв'язку за схемою А-Т, Г-Ц, Т-А, Ц-Г ( рис. 9.28). Навпроти кожного нуклеотиду однієї з батьківських ланцюгів ДНК розташовується комплементарний нуклеотид синтезується ДНК. В результаті утворюється ланцюжок ДНК являє собою точну копію вихідної ланцюга.
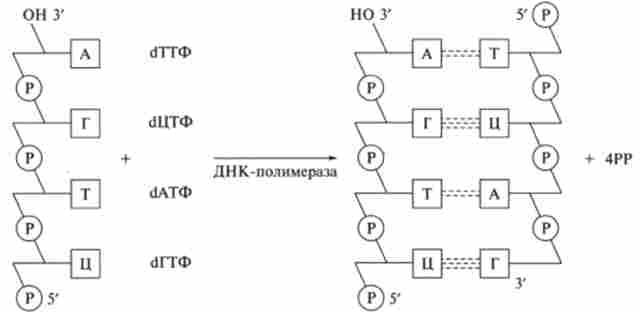
Мал. 9.28. Полімеризація дочірньої ланцюга ДНК за принципом комплементарності
Полімерази в ході транскрипції пов'язують приєднані нуклеотиди в нову ланцюг ДНК.
Ініціація біосинтезу дочірніх ланцюгів ДНК вимагає попереднього синтезу на матриці ДНК затравочного олігорібонуклеогіда - праймера. Праймер має вільну гідроксильну групу рибози. Цей коротколанцюговий олігорі- бонуклеотід синтезується комплементарно на матриці ДНК за участю ферменту ПРАЙМАЗИ з полімеразної активністю.
З кінцевого гідроксилу рибози праймера починається синтез дочірньої ланцюга ДНК. Синтез здійснюється в результаті реакції етерифікації між ОН-групою кінцевого рибонуклеотиду праймера і ОН-групою фосфату першого де- зоксірібонуклеотіда Nui в суворій відповідності з принципом комілементарно- сти (рис. 9.29). При цьому звільняється пирофосфат РР ,.
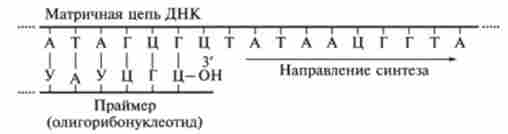
Мал. 9.29. Синтез дочірньої ланцюга ДНК з кінцевого гідроксилу рибози праймера
Надалі цей фрагмент РНК, комплементарно приєднаний до новоствореної ланцюга ДНК, руйнується під дією ДНК-полімерази I і виникла пролом забудовується олігодезоксирибонуклеотиди за допомогою тієї ж ДНК-полімерази I. Цілком припустимо припущення, що синтез праймера з олігорібонуклеотіда має глибокий біологічний сенс, оскільки в цьому випадку можуть усуватися помилки, що неминуче виникають при ініціації реплікації ДНК.
Запропоновано різні механізми біосинтезу ДНК за участю відомих ферментів і білкових факторів. Вони грунтуються головним чином на даних, отриманих в дослідах in vitro (В колбі).
На основі вивчення кишкової палички Е. coli в механізмі синтезу ДНК виділяють ініціацію (початок), елонгацію (продовження) і терминацию (припинення) синтезу. Кожна з цих стадій вимагає участі специфічних ферментів і білкових факторів.
ініціація є початком синтезу дочірніх нуклеотидних ланцюгів ДНК. У ній бере участь як мінімум вісім добре вивчених ферментів і білків. Ініціації передує синтез праймера на матриці ДНК. В ході ініціації до ланцюгів ДНК послідовно приєднуються ДНК-раскручівающіс і ДНК-зв'язуючі білки, а потім комплекси ДНК-полімерази і праймаза (рис. 9.29).
Ініціація представляється єдиною стадією реплікації ДНК, яка точно регулюється. Однак її деталі не розкриті і інтенсивно досліджуються.
елонгація синтезу ДНК включає два різних за механізмом процесу синтезу лідируючій і відстає ланцюгів на розплетених батьківських ланцюгах ДНК. Лідируюча ланцюг починається з синтезу праймера (за участю ПРАЙМАЗИ) в точці початку реплікації. Потім до праймеру приєднуються дезоксирибонуклеотидів під дією ДНК-полімерази III. Далі синтез протікає безперервно, дотримуючись за переміщенням вилки реплікації.
Синтез відстає ланцюга на другий ланцюга Дик протікає в напрямку, протилежному руху вилки реплікації, і починається фрагментарно. Фрагменти синтезуються окремо, починаючи з синтезу праймера. Праймер може переноситися з готового фрагмента в точку старту біосинтезу подальшого фрагмента за допомогою одного з білкових факторів реплікації. Елонгація завершується відділенням олігорібонуклеотідних праймерів, об'єднанням окремих фрагментів ДНК за допомогою ДНК-лігази і формуванням дочірньої ланцюга ДНК.
Передбачається, що може здійснюватися пов'язаний і узгоджений механізм синтезу лідируючій і відстає ланцюгів ДНК за участю полимераз і всього комплексу праймаза.
Терминация синтезу ДНК настає, коли ДНК-матриця пройдена до кінця.
Точність реплікації ДНК надзвичайно висока. Можлива одна помилка на 1000 ланок ланцюга, однак і ця помилка зазвичай виправляється за рахунок процесів репарації. Правильність реплікації забезпечується принципом комплементарності і активністю ДНК-полімерази, здатної розпізнати і виправити помилку.
Принцип комплементарності, на основі якого побудована двухспіральная молекула ДНК, діє і при синтезі мРНК. Гак само як і при синтезі ДНК, з кожним нуклеотидом однієї з ланцюгів ДНК зв'язується водневими евзямі комплементарний нуклеотид іРНК. Слід зазначити, що в іРНК замість нуклеотиду Т присутній нуклеотид У. Таким чином, навпаки Гднк розташовується Црнк »навпроти Цднк - ГЖК, навпаки Аднк - Урнк, навпаки ТДНдо - Атк.
В результаті синтезована ланцюг іРНК являє собою точну копію ланцюга ДНК. Таким шляхом інформація, що міститься в гені, переписується на іРНК. Тому цей процес і назвали транскрипцією (лат. «Transcriptio» - переписування).
По завершенні транскрипції молекули іРНК переміщуються до місця синтезу білка, т. Е. До рибосом. Туди ж з цитоплазми надходять амінокислоти, з яких будується білок. Кожна амінокислота переноситься в рибосому спеціалізованої транспортної РНК (тРНК). Так як до складу природних білків входить 20 різних амінокислот, очевидно, існує не менше 20 різних тРНК.
У ланцюзі тРНК є 4-7 послідовних нуклеотидних ланок, комплементарних один одному. На рис. 9.30 вони позначені буквами А Б В Г. У цих ділянках між комплементарними нуклеотидами утворюються водневі зв'язку.
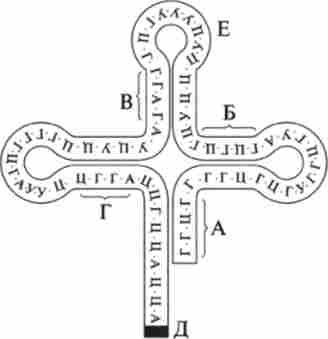
Мал. 9.30. структура тРНК
генетичний код
|
перше підставу |
друга підстава |
третє підставу |
|||
|
У (А) |
Ц (Г) |
А (Т) |
Г (Ц) |
||
|
У (А) |
фен |
сер |
тир |
цис |
У (А) |
|
фен |
сер |
тир |
цис |
Ц (Г) |
|
|
лей |
сер |
- |
- |
А (Т) |
|
|
лей |
сер |
- |
Три |
Г (Ц) |
|
|
Ц (Г) |
лей |
про |
Гіс |
Apr |
У (А) |
|
лей |
про |
Гіс |
Apr |
Ц (Г) |
|
|
лей |
про |
Глн |
Apr |
А (Т) |
|
|
лей |
про |
Глн |
Apr |
Г (Ц) |
|
|
А (Т) |
Ілі |
Тре |
Асі |
сер |
У (А) |
|
Ілі |
Тре |
Асі |
сер |
Ц (Г) |
|
|
Ілі |
Тре |
Ліз |
Apr |
А (Т) |
|
|
мет |
Тре |
Ліз |
Apr |
Г (Ц) |
|
|
Г (Ц) |
Вал |
Ала |
асп |
гли |
У (А) |
|
Вал |
Ала |
асп |
попелиці |
Ц (Г) |
|
|
Вал |
Ала |
гли |
гли |
А (Т) |
|
|
Вал |
Ала |
гли |
Г'лі |
Г (Ц) |
|
Таблиця 9.3 в результаті виникає складна петлистая структура, схожа за формою на листок конюшини. У верхній частині молекули тРНК (рис. 9.30, Е) розташований триплет нуклеотидів, який за генетичним кодом відповідає певній амінокислоті. Цей триплет називають кодовою кодоном.
У ніжки «листка конюшини» (рис. 9.30, Д) знаходиться ділянка, що зв'язує амінокислоту. Нуклеотидний склад кодових триплетів тРНК комплементарен нуклеотидному складу триплетів і PH К.
Наприклад, у аланиновой тРНК кодовий триплет ЦГА (в іРНК йому комплементарен триплет ГЦУ). у валіновой тРНК кодовий триплет ЦАА (в іРНК йому комплементарен триплет ГУУ) (табл. 9.3). У тРНК, зображеної на малюнку, кодовий триплет УУУ, відповідний амінокислоті лізину, - лізіновая тРНК. Вона приєднує і транспортує в рибосому амінокислоту лізин.
У табл. 9.3 наведено склад триплетів, якими закодовані всі 20 амінокислот (назви амінокислот скорочені).
Користуватися таблицею просто. Перший нуклеотид в триплеті береться з лівого вертикального ряду, другий - з верхнього горизонтального і третій - з правого вертикального. Там, де перетнуться лінії, що йдуть від всіх трьох нуклеотидів, і знаходиться шукана амінокислота. Припустимо, потрібно дізнатися, про яку амінокислоті несе інформацію триплет УГГ в іРНК. Зліва по вертикалі беремо У, зверху - Г, праворуч по вертикалі - Г. Лінії перетинаються на «Три», т. Е. Триптофан. У ДНК ця амінокислота закодована кодонів АЦЦ.
Синтез дочірньої молекули дезоксирибонуклеїнової кислоти відбувається в процесі поділу клітини на матриці батьківської молекули ДНК. При цьому генетичний матеріал, зашифрований в ДНК, подвоюється і ділиться між дочірніми клітинами. У клітинному циклі час синтезу і подвоєння ДНК називається S-стадією. За нею йдуть стадії G2 і М - мітоз (поділ клітин).