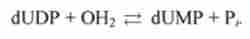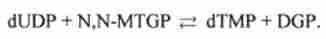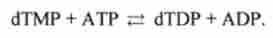Будівельні блоки ДНК, т. Е. Дезоксі- рибонуклеотиди, утворюються з рибонуклеотидов в реакції відновлення ри- бонуклеотіда до відповідного дезоксіпроізводного.
Всі чотири рібонуклеозіддіфосфата відновлюються в дезоксірібонукле- озіддіфосфати dADP, dGDP, dCDP. dUDP за участю складної ферментної системи, що складається щонайменше з чотирьох різних ферментів.
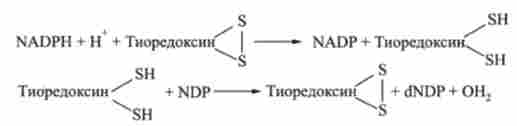
Сенс перетворення рибонуклеотидов в дезоксирибонуклеотидів зводиться до відновлення рибози в 2-дезоксирибози, що вимагає наявності двох атомів водню. Їх джерелом є відновлений нікотінамідаденіннуклео- тідфосфат NADPH. У передачі атомів водню бере участь термостабільний білок - тіоредоксін, що містить дві вільні SH-групи, за допомогою яких і відбувається перенесення водневих атомів від N А ПРИ до рібонуклеозіддіфосфату. Тіоредоксін легко окислюється, перетворюючись в дисульфидную S-S-форму. Його відновником служить фермент тіоредоксінредуктази, що містить окислений флавінаденіндінуклеотід PAD. Потім відновлений тіоредоксін відновлює нуклеозіддіфосфат NDP до дезоксірібонуклеозіддіфосфата dNDP:
Реакція каталізується рибонуклеотидредуктазу.
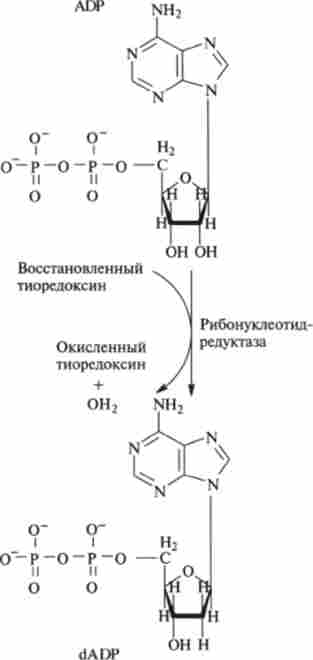
На рис. 9.26 перетворення рибонуклеотидов в дезоксирибонуклеотидів показані на прикладі нуклеотидів - похідних аденіну ADP -> dADP.
Мал. 9.26.Біосинтез дезоксірібонуклеотідов - дезоксіаденозілдіфосфата dADP з рибонуклеотиду аденозілдіфосфата ADP
У ДНК містяться залишки тимідилової кислоти dTMP замість залишків урі- ДИЛОВА кислоти UMP, які присутні в РНК. Шлях синтезу dTMP починається з освіти з уридилдифосфата UDP дезоксіуріді л дифосфата dUDP. Потім dUDP гідролізується з утворенням дезоксіуріділмонофосфата:
Потім dUMP в результаті реакції метилування за допомогою N, N- метілентетрагідрофолата MTGP перетворюється в дезоксітіміділат dTMP (реакція каталізується тіміділатсінтетазу). Другим продуктом реакції служить д і гідрофолат DGP:
Фосфорилювання dTMP призводить до утворення dTDP:
Синтез дезоксирибонуклеозид-5-трифосфату завершується в результаті наступних реакцій, які каталізують відповідними кіназами:
Будова типовою клітини багатоклітинного організму - біологія. Частина 1 Клітини багатоклітинних організмів, як тварин, так і рослинних, відокремлені від свого оточення оболонкою. Клітинна оболонка, або плазмалемма, тваринних клітин утворена мембраною, покритий зовні шаром гликокаликса товщиною 10-20 нм. Основними складовими гликокаликса служать комплекси полісахаридів
Будова серця - вікова анатомія і фізіологія. Т.2 опорно-рухова і вісцеральні системи 4 р діафрагма Мал. 9.2. Положення серця в грудній порожнині Серце розташоване в грудній порожнині між легкими і зазвичай зміщений вліво (рис. 9.2). Серце розташовується асиметрично: його вісь спрямована праворуч-зверху, вперед-вниз і вліво. Своїм підставою серце звернено до хребта, а сто верхівка
Будова молекул ДНК і РНК - генетика в 2 Ч. Частина 1 Перший доказ ролі ДНК як носія спадкової інформації організмів привернув величезну увагу до вивчення нуклеїнових кислот. У 1869 р Ф. Мішер виділив з ядер клітин особливу речовину, яку назвав нуклєїнах. Через 20 років ця назва була замінена терміном нуклеїнова кислота. У 1924 р Р. Фельгена
Біосинтез замінних амінокислот - біохімія частина 2. Людина і тварини здатні синтезувати тільки 10 з 20 амінокислот, необхідних для синтезу білка, - це замінні амінокислоти (24.2). Шляхи біосинтезу цих амінокислот різноманітні, але при цьому вони володіють однією важливою властивістю: (Т) вуглецевий скелет амінокислот утворюється з проміжних
Біосинтез піримідинових рибонуклеотидів - біохімія частина 2. Застосування мічених атомів дозволило з'ясувати походження кожного з атомів піримідинового кільця: Біосинтез урідінмонофосфата (УМФ) - загального попередника всіх піримідинових нуклеотидів включає шість реакцій. 1. Освіта карбамоілфосфата при дії карбамоілфосфатсінте- тази II (КФС II). Донором
Біосинтез фосфогліцерідов - біохімія людини Найбільш важливі фосфогліцерідов є компонентами мембран, а також ліпопротеїдів, що виконують транспортну функцію. Ці фосфогліцерідов утворюються по розгалуженому метаболічного шляху, що починається з фосфатидного кислоти Реакції шляхів біосинтезу фосфогліцерідов (рис. 9.18) локалізовані головним