| Головна |
| «« | ЗМІСТ | »» |
|---|
Ембріологія (від гр. Embryon - зародок, logos - вчення) - наука про розвиток зародка, яка вивчає індивідуальний розвиток тварин з моменту зародження (запліднення яйцеклітини) до народження.
Ембріогенез - це складно протікає процес, в якому знайшли відображення еволюція хребетних (філогенез) та індивідуальний розвиток (онтогенез).
Ембріологія розглядає розвиток і будова статевих клітин (гаметогенезу) і основні етапи ембріогенезу: запліднення, дроблення, гаструляції, закладку осьових органів і органогенез, розвиток провізорних (тимчасових) органів.
Мейоз (від гр. Meiosis - зменшення) - тип поділу статевих клітин - гамет.
Мейоз виник разом з появою статевого запліднення, при якому зливаються чоловіча і жіноча гамети і число хромосом подвоюється - диплоїдний набір (від гр. Diploos- подвійний). Оскільки статеве розмноження повторюється з покоління в покоління, число хромосом в ядрі мало б подвоюватися нескінченно, проте цього не відбувається, так як під час мейозу зменшується число хромосом з диплоїдного до гаплоїдного (від гр. Gaploos - одинарний).
Мейоз - єдиний, безперервний процес, що складається з інтерфази і двох послідовних поділів (мейоз I і мейоз II); кожне з поділів складається з тих же фаз, що і мітоз (профази, метафаза, анафаза, телофаза).
Интерфаза протікає так само, як і при мітозі: збільшується число органоїдів і відбувається редуплікація ДНК (кожна хромосома представлена парою хроматид, з'єднаних центромерой).
М е й о з I - перший мейотичне розподіл називають редукційним (від лат. Reduction - зменшення), так як з диплоїдних клітин (2п2с) утворюються гаплоїдні клітини (п2с).
В профазу I компактно укладається генетичний матеріал - спирализуются хромосоми. До протилежних полюсів клітини переміщаються центриоли, і між ними формується ахроматі- нове веретено, що складається з безлічі микротрубочек, т. Е. Такі ж зміни, як в профазі мітозу.
На відміну від мітозу профази I триваліша, і в ній виділяють кілька стадій: лептотене, зіготене, пахітене, діплотене, діакінеза (рис. 9).
Лептотена (від лат. Leptos - тонкий, taenia - стрічка, нитка) - рання стадія, в якій ядерний матеріал представлений у вигляді довгих і тонких ниток.
Зиготена (від rp. Zygoo - з'єднувати), або синаптична стадія (від лат. Synapsis - з'єднання), характеризується початком кон'югації гомологічних хромосом, які об'єднуються сінаптонемальним комплексом в біваленте. Гомологічні хромосоми (від гр. Gomologos - відповідний) зближуються і з'єднуються в декількох точках - кон'югація. Кожна така пара (одна материнська, інша батьківська) хромосом утворює пари - діади, що складаються з чотирьох хроматид, і їх називають бівалентом. Між гомологічними хромосомами здійснюється перехрещення з обміном відповідними ділянками; гени з однієї хромосоми виявляються пов'язаними з генами іншого хромосоми, такий обмін однаковими ділянками (генами) називають кросинговером (від лат. crossing - схрещування, over - через).
Пахитена (від лат. Pachys - товстий) - стадія, в якій пари гомологічних хромосом перекручуються один з одним, спирализуются і товщають.
Диплотена (від лат. Diploos - подвійний) - процес формування зошити. У кожній парі хромосом виникає поздовжня щілину, і в двох кон'югованих хромосомах утворюються чотири хрома- тіди. Між гомологічними хромосомами виникають сили відштовхування, і вони починають віддалятися один від одного в першу чергу в області центромер, але залишаються пов'язаними в ділянках, де стався кросинговер, - хіазмі.
Діакінеза - завершальна стадія профази I, в якій гомологічні хромосоми утримуються разом лише в окремих точках - хіазмі, набуваючи химерну форму кілець, хрестів, вісімок.
В метафазу I гомологічні хромосоми розташовуються в екваторіальній площині (посередині) клітини і кожна з них складається з двох хроматид.
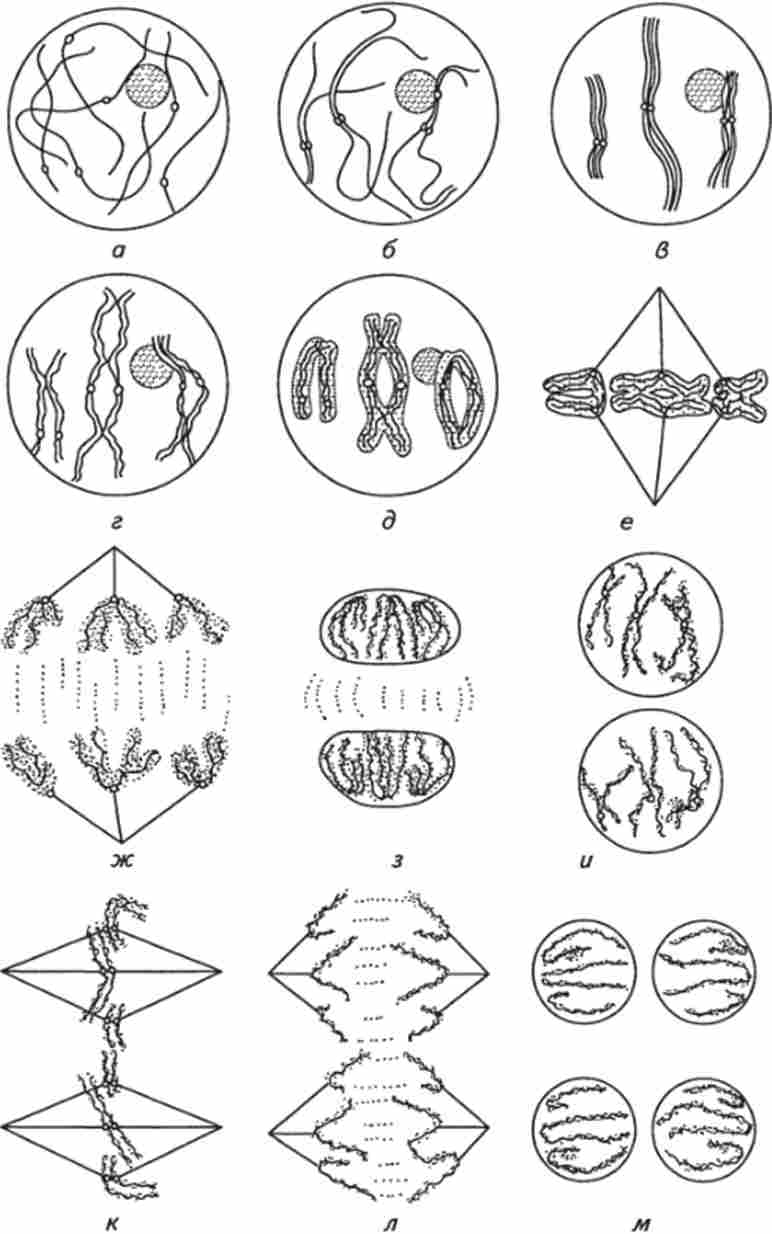
Мал. 9. Мейоз в тваринній клітині:
а - Лептотена; б - Зиготена; в - Пахітена; г - діплотена; д - діакінеза; е - метафаза I; ж - анафаза I; з - телофаза I; і - профази II; до - мстафаза II; л - анафаза II; м - телофаза II
В анафазу / Скорочуються нитки ахроматинового веретена захоплюють кожну з гомологічниххромосом до протилежних полюсів клітини.
В телофазу I хромосоми досягають полюсів клітини, розкручуються (деспирализуются), стають погано помітними; формуються ядерце і ядерна оболонка, в результаті оформляються два ядра, одночасно з цим відбувається поділ цитоплазми - цитокинез.
М е й о з II - друга мейотичне розподіл називають екваці- онним (тільки що утворилися дві клітини діляться).
В профазу II хроматиди спирализуются, ядерце і ядерна оболонка стають невидимими. До протилежних полюсів клітини переміщаються центриоли, і між ними формується ах- роматіновое веретено, що складається з безлічі микротрубочек.
В метафазу II хромосоми розташовуються в екваторіальній площині (посередині) клітини (число хромосом в 2 рази менше, ніж в соматичної клітці).
В анафазу II скорочуються нитки ахроматинового веретена захоплюють кожну з хроматид до протилежних полюсів клітини.
В телофазу // хроматиди досягають полюсів клітини, розкручуються (деспирализуются), стають ледь помітними; формуються ядерце і ядерна оболонка, в результаті оформляються два ядра, одночасно з цим відбувається цитокинез.
Таким чином, з вихідної батьківської клітини з диплоїдним набором утворюються чотири дочірні клітини з гаплоїдний набором хромосом. У процесі запліднення статеві клітини зливаються і диплоїдний набір хромосом відновлюється.
При мейозі гомологічні хромосоми з'єднуються в пари, потім в результаті першого мейотичного поділу розходяться по одній в дочірні клітини. Під час другого мейотичного поділу гомологічні хромосоми розходяться в нові дочірні клітини.
Механізм мейозу забезпечує сталість числа хромосом. Якби не відбувалося редукції числа хромосом при гаметоге- незе, то з покоління в покоління зростало б число хромосом і був би втрачений один з суттєвих ознак кожного виду - постійність числа хромосом.
При мейозі утворюються нові комбінації хромосом. Так, в диплоїдний набір хромосоми мають подвійне походження: у кожній гомологичной парі одна з хромосом від батька, інша-від матері. У сперміях і яйцеклітинах утворюються нові поєднання спадкової інформації.
У процесі кросинговеру також відбувається рекомбінація генетичного матеріалу. Практично всі хромосоми, що потрапляють в гамети, мають ділянки, що відбуваються як від спочатку батьківських, так і від спочатку материнських хромосом.
Цим досягається ще більша ступінь перекомбінації спадкового матеріалу. У цьому одна з причин мінливості організму, що дає матеріал для відбору.
Таким чином, принципова відмінність статевого розмноження від безстатевого полягає в тому, що воно призводить до величезної мінливості, утворення форм з новими спадковими властивостями в результаті перекомбінації різних властивостей обох батьків, в той час як при безстатевому розмноженні дочірні організми повторюють спадкову інформацію єдиною батьківської особини.