| Головна |
| «« | ЗМІСТ | »» |
|---|
Одна з головних характеристик типу хордові - це наявність трубчастої НС. Вона розвивається з ектодермальну нервової трубки і займає в організмі дорсальне положення, т. Е. Централізація і концентрація нейронів у хордових пов'язана зі спинний стороною тіла, а не з черевної (черевна нервова ланцюжок), як у більшості безхребетних.
Загальний план будови НС у всіх хордових однаковий - це нервова трубка, від якої відходять парні нерви. У нижчих хордових НС влаштована досить просто. Розглянемо як приклад НС примітивного хордової тварини ланцетника (підтип безчерепні).
ланцетник - морська тварина, що веде вкрай малорухливий спосіб життя і має дуже примітивні органи чуття (ні зоровим, ні слуховим апаратом ланцетник не має). З боків тіла у нього лежать м'язи, розташовані правильними, що повторюють один одного сегментами (міомеров). Кісткового скелета, як і черепа, немає. Скелет представлений хордою (спинний струною), яка зберігається і в підтип хребетні (по крайней мере, в початкові періоди ембріогенезу).
ЦНС ланцетника (рис. 16.8) лежить над хордою, не досягаючи її переднього кінця.
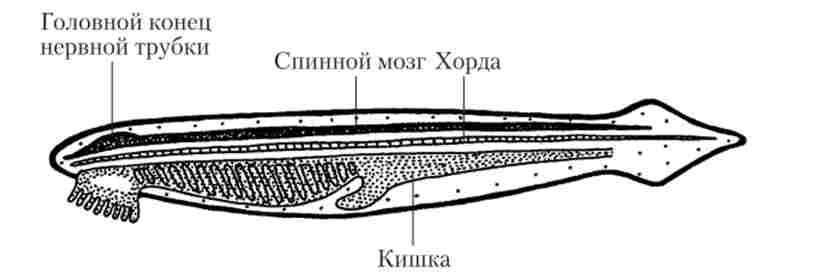
Мал. 16.8. Нервова система ланцетника
В цілому у ланцетника тільки намічається диференціювання ЦНС на ГМ і СМ. Проявляється це в тому, що спинномозковий канал на передньому кінці тіла трохи розширюється, утворюючи один недиференційований мозкової шлуночок. Від переднього кінця відходять дві пари головних нервів - нюхових і термінальних. Від спинного мозку відходять метамерно розташовані спинномозкові нерви. При цьому спинні і вентральні корінці не об'єднує, а являють собою самостійні нерви. Мієлінізація нервових волокон відсутня.
В ряду хребетних еволюція НС головним чином пов'язана з усе більшою енцефалізаціі (encephalon - ГМ), т. Е. Дифференцировкой і збільшенням її рострального кінця з переміщенням туди деяких функцій і систем інтеграції. У всіх хребетних ГМ проходить в своєму онтогенезі стадію трьох, а потім і п'яти мозкових міхурів. І у всіх дефінітивний ГМ складається з п'яти відділів: кінцевого і проміжного мозку (передній мозок), середнього мозку і ромбовидного мозку з мозочком.
Перш ніж розглянути філогенез цих частин головного мозку, згадаємо основні класи хребетних.
клас круглороті (Міноги і міксин) - найпримітивніші хребетні, що входять в розділ безщелепні (іноді міног і міксин виділяють в окремі класи). Це водні тварини, провідні полупаразі- тичний спосіб життя. Від риб вони відрізняються відсутністю щелеп і парних кінцівок, непарним органом нюху. У круглоротих, як і у риб, є специфічний орган почуттів - бічна лінія. У внутрішньому вусі присутні тільки два (а не три, як у всіх інших хребетних) півколових каналу.
надклас риби включає два великі класи - хрящові і кісткові, в організації яких є принципові відмінності. Але в порівнянні з круглоротими все риби виявляють прогресивні риси розвитку. Риби мають парні кінцівки (плавники), що дозволяють здійснювати більш складні рухи, і вони ведуть більш активний спосіб життя. У риб вже є всі типові для хребетних органи чуття.
клас амфібії (Земноводні) - це перші хребетні, що вийшли в процесі еволюції на сушу. Однак вони не повністю відірвалися від водного середовища - розмножуються амфібії в воді, причому в воді тільки прісної, що дуже обмежує їх поширення. Найбільш відомі амфібії - це безхвості жаби, хвостаті тритони і саламандри. Земноводні мають типову для всіх наземних тварин п'ятипалу кінцівку ричаговими типу. Однак руху у них досить примітивні - передні кінцівки не мають зв'язку з осьовим скелетом, що дуже обмежує їх рухливість.
Всі перераховані вище хребетні тварини належать до групі анампій - первічноводних. Вони або живуть у воді, або, як амфібії, хоча б в ній розмножуються.
Наступних за ними в еволюційному розвитку плазунів, птахів і ссавців відносять до амніоти - первічноназемним, які вже повністю відірвалися від води і на суші не тільки живуть, а й розмножуються. Деякі амніоти, так звані повторноводних, в процесі еволюційного розвитку знову перейшли до життя у воді, але розмножуються вони все-таки на суші (крокодили, водні черепахи, тюлені і т. П.). Єдиний виняток - китоподібні (кити, дельфіни), які повністю втратили зв'язок із сушею. Незалежність в своєму розмноженні від води забезпечила амніоти широке поширення по земній кулі. Вся їхня організація пов'язана з наземним способом життя. Це проявляється в більшій диференціювання всіх відділів тіла і всіх систем органів. Удосконалення опорно-рухової системи значно збільшує рухливість амниот в порівнянні з анамнії. І відносні розміри головного мозку, особливо півкуль і мозочка, у амниот зростають. Всі ці зміни забезпечують амніоти вищий рівень життєдіяльності, кращу адаптацію до змін навколишнього середовища, більш складну вищу нервову діяльність. У зв'язку з усім вищевикладеним анамній прийнято називати нижчими хребетними, а амниот - вищими.
рептилії (Плазуни) - перший клас первічноназемних тварин. На відміну від амфібій, що мають голу слизову шкіру, у плазунів шкіра суха, покрита лусками або щитками. Це ящірки, змії, крокодили, черепахи.
класи птахів і ссавців принципово відрізняються від всіх інших тварин теплокровностью - здатністю підтримувати постійну температуру тіла. Це дозволяє їм в найменшій мірі залежати від навколишнього середовища і вести набагато більш активний спосіб життя.
Філогенез основних відділів головного мозку (рис. 16.9). Відразу треба відзначити, що чим ростральними розташований відділ мозку, тим більші зміни в філогенезі зазнає цей відділ.
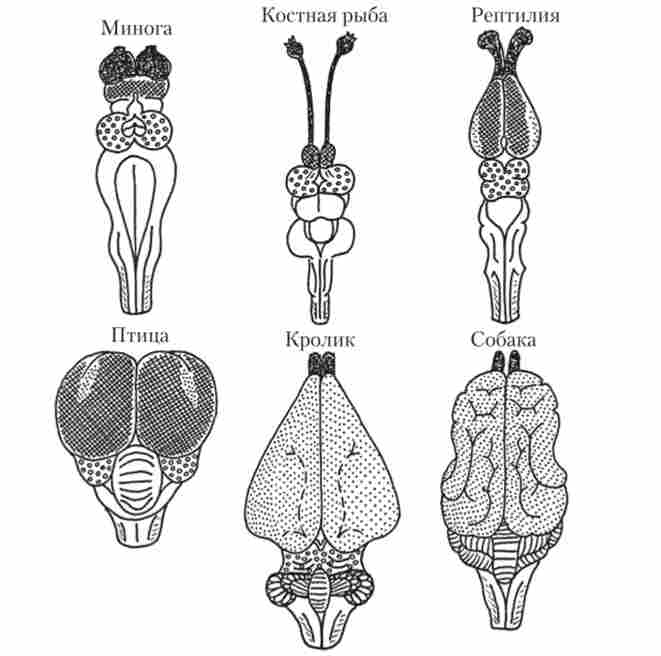
Мал. 16.9. Головний мозок різних класів хребетних з дорсальній боку:
У всіх хребетних в стовбурі мозку знаходяться сенсорні і рухові центри, ядра РФ, а також ядра всіх черепних нервів, за винятком зорового, нюхового і кінцевого. Диференціація ядер і їх число наростають в ході філогенезу. З усіх структур стовбура найбільш чіткі зміни в процесі філогенезу простежуються на рівні середнього мозку, який в своєму розвитку в першу чергу пов'язаний із зоровою функцією. Середній мозок складається з верхньої частини - ТЕКТУМ (даху) і нижньої - тегментума (покришки). У вищих хребетних йод покришкою формується базис (основу), що складається з тих, які сходять трактів, що йдуть від кінцевого мозку в нижележащие структури. У анамній середній мозок - це сама виражена частина головного мозку, яка розглядається як вищий інтеграційний центр.
Основна частина ТЕКТУМ всіх хребетних, за винятком ссавців, - це парні великі зорові частки (верхні горбки у ссавців), величина і розвиток яких корелює зі значенням зорової сенсорної системи для даного виду. У ссавців і рептилій до верхніх горбках додаються нижні, слухові, в результаті чого формується четверохолмие. У цих тварин співвідношення між верхніми і нижніми горбками може служити показником провідного значення зорового або слухового аналізаторів. Наприклад, у копитних, деяких хижаків, приматів верхні горбки розвинені більше, ніж нижні; у китоподібних і кажанів - навпаки. Треба додати, що зорові горбки ссавців зазвичай розвинені слабше, ніж у більш примітивних тварин, що, мабуть, пов'язано з тим, що у них з'являються добре розвинені коркові і таламичні зорові центри. У всіх хребетних зорові горбки мають шарувату організацію, в якій чергуються клітинні і волокнисті шари, причому кількість шарів нерідко набагато більше, ніж у ссавців. Так, в ТЕКТУМ деяких рептилій налічують до 14, а птахів - до 15 шарів.
У круглоротих мозочок дуже маленький, макроскопично майже невиразні. Він являє собою поперечний валик, що йде але переднього краю ромбовидноїямки. Його виникнення пов'язують з появою внутрішнього вуха з його вестибулярним апаратом і з розвитком органів бічної лінії. У мозочку круглоротих вже присутні зернисті клітини і більші клітини - попередники клітин Пур- кінье. В ході подальшої еволюції мозочок збільшується в розмірах, його поверхня покривається борознами, внаслідок чого стає можливим говорити про частки мозочка. Поступово частина нейронів мозочка переходить на його поверхню, і в амніот ці клітини утворюють кору мозочка. Вже у риб в товщі мозочка виділяються клітини, аксони яких стають ефферентов мозочка. Їх кількість зростає, і в амніот вони об'єднуються в ядра.
Мозочок у риб розвинений набагато краще, ніж у круглоротих, що в першу чергу пов'язано з більш активними рухами цієї групи тварин. Проте у різних представників риб може спостерігатися дуже велика різниця в розмірі мозочка, що багато в чому пов'язано з їхнім способом життя. Так, наприклад, у Двоякодихаючих і кистеперих риб, що живуть в заростях на дні водойм, мозочок дуже невеликий. Мозочок у більшості амфібій розвинений дуже слабо (його можна порівняти з мозочком Дводишні риб), що відображає простоту моторики тварин цього класу.
В цілому мозочок у анамній зазвичай складається з центральної частини - тіла і бічних виростів, які називають аурикула або ушковидная частками мозочка. Аурикула, мабуть, пов'язані з вестибулярним апаратом і органами бічної лінії. У амниот аурикула перетворюються в Клочкова-узелковую (флокули-нодулярну) частку, яка пов'язана з вестибулярними ядрами (бокової лінії у амниот немає).
У вищих хребетних мозочок розвинений в набагато більшому ступені, ніж у анамній. Поверхня має виражені борозни і звивини, в глибині мозочка лежать ядра. Однак у рептилій і птахів мозок складається тільки з хробака; півкулі практично відсутні. Є лише невеликі бічні випинання флокули-нодулярної частки. І тільки у ссавців з'являються справжні півкулі мозочка і формується неоцеребеллум.
У всіх хребетних в проміжному мозку виділяють таламус (зоровий бугор), гіпоталамус і епіталамус, причому кожен з відділів може бути виражений в більшому або меншому ступені. Субталамічні освіти диференціюються тільки у ссавців.
Найбільші структурні зміни в ряду хребетних зазнає таламус, що, мабуть, обумовлено його взаємними зв'язками з інтенсивно розвиваються відділами кінцевого мозку. Основний напрямок перетворень таламуса пов'язано зі збільшенням його маси (у ссавців таламус - найбільше освіту діенцефалона), а також з усе більшою диференціацією його ядер.
При порівнянні структур гіпоталамуса у різних класів хребетних його зазвичай ділять на перивентрикулярна (околожелудочкового), медіальну і латеральну зони. І хоча в гіпоталамусі практично всіх хребетних виділяють області, які виконують подібні функції, ступінь розвитку різних зон різна. У більшості анамній основна маса клітин знаходиться в перивентрикулярной зоні, у амниот - в медіальній. Причому у рептилій і деяких амниот найбільш диференційовані структури медіальної зони, а у ссавців і птахів - латеральної. Загальна риса гіпоталамуса всіх хребетних - присутність в перивентрикулярной зоні нейросекреторну клітин, число яких особливо велике у анамній в преоптичної області, а в амніот - в паравентрикулярного і преопті- зації ядрах. У медіальній зоні у всіх досліджених хребетних знаходять супрахіазменние ядро, яке отримує зорові афференти.
Як відомо, до складу епіталямуса ссавців входять ядра повідків (хабенулярние ядра) і заліза внутрішньої секреції - епіфіз. Ядра повідків з'являються вже у круглоротих і надалі присутні у всіх інших хребетних. Що стосується епіфіза, то у нижчих хребетних він виконує інші функції, і в цьому випадку його прийнято називати пинеальная органом. Ростральні від нього у анамній є пара- пинеальная, або тім'яної, орган (тім'яної очей). Обидва цих освіти містять фоторецепторні клітини, які сприймають інтенсивність світла, але не дають зображення. Деякі тварини мають обома цими органами, у інших один з органів є переважаючим. У вищих хребетних пинеальная орган втрачає світлочутливі клітини і перетворюється в епіфіз. Тім'яної очей у деяких рептилій (гаттерия, ящірки) залишається, а у птахів і ссавців редукується.
У вищих хребетних кінцевий мозок найбільш складно влаштований відділ мозку, але основні його структури простежуються і у анамній. Назви відділів кінцевого мозку в ряду хребетних дещо відрізняються від вже відомих нам по попереднім головам. Це пов'язано з необхідністю порівняльно-анатомічного аналізу мозку тварин, що знаходяться на різних щаблях еволюційної драбини, причому анатомічна номенклатура в різних класах хребетних не завжди однакова. Як правило, привласнення найменування відділам кінцевого мозку проводиться з урахуванням його ембріонального розвитку, цитоархітектоніки, міелоархітектоніка і як аферентних, так і еферентних зв'язків з підкірковими структурами. Викладемо (з деякими спрощеннями) сучасну номенклатуру кінцевого мозку.
Нагадаємо, що кінцевий мозок у ссавців складається з знаходиться зверху кори великих півкуль і розташованих в товщі півкуль базальних ядер. У всіх хребетних кінцевий мозок формується з крилоподібні пластинки (див. Параграф 4.1). Її клітини поділяються на дорсальну і вентральную зони. Дорсальная зона утворює поверхню півкуль - плащ, або палліум (pallium)] вентральна - формації суб палліум - базальні ядра. У палліум виділяють pallium lateralis, pallium medialis, pallium dorsalis. У амниот з них формуються відповідно палеокортекс, архикортекс і неокортекс. Для анамній поняття «палліум» не рівнозначно поняттю «кора великих півкуль». Палліум - це фактично дах кінцевого мозку, ті тканини, які утворюють його поверхню і далеко не завжди мають корковую організацію. У субпалліуме виділяють неостріатум (хвостате ядро і шкаралупа), палеостріатум (блідий кулю), архістріатум (мигдалеподібної комплекс) і септума.
Стародавні хребетні отримували інформацію про навколишній світ головним чином за допомогою нюху. І розвиток кінцевого мозку в першу чергу пов'язано з нюхової сенсорної системою. Майже у всіх хребетних ростральна частина переднього мозку випинається, утворюючи нюхові цибулини, мають загальний план будови у всіх класів. Найчастіше цибулини безпосередньо пов'язані з півкулями, однак у деяких хребетних, в тому числі і у приматів, вони винесені на якусь відстань і з'єднані з мозком нюхових трактом. Але навіть у круглоротих кінцевий мозок виконує не тільки нюхову функцію. Туди приходять афференти від інших сенсорних систем, і в якійсь мірі навіть при низькому рівні розвитку кінцевого мозку притаманні інтегрують функції.
У круглоротих дах півкуль позбавлена нервової тканини, вона тонка, епітеліальна. Основна маса нейронів зосереджена навколо бічних шлуночків. У риб півкулі розвинені вже набагато краще, але на їх поверхні нервові клітини відсутні, вони лежать глибше. У багатьох хрящових риб вперше в ряду хребетних формується коркова пластинка - шар клітин, відокремлених від нерівентрікулярной зони. У амфібій кінцевий мозок стає набагато більше, ніж у більшості риб. У ньому в зародковому вигляді представлені всі основні формації. У даху мозку з'являється нервова тканина, особливо розвинений медіальний палліум - архіпал- ліум (зачаток гіпокампу) - в цьому відділі нейрони займають всю товщу стінки мозку, і він відокремлюється від субпалліума прошарком білої речовини. У латеральному палліум простежуються нюхові центри, які в подальшому (у амниот) дадуть палеокортекс. В базальних гангліях можна виділити відділи, подібні до нео-, палео-і архістріатумом амниот.
Починаючи з рептилій, кінцевий мозок стає найбільшою частиною ЦНС. В останні роки уявлення про його будову у рептилій і птахів піддані значному перегляду. Оскільки у цих тварин основна маса кінцевого мозку має ядерну будову, їх півкулі довгий час відносили до субпалліальним формаціям. Однак завдяки сучасним (перш за все - ембріологічним) методам з'ясувалося, що більшість відділів, які вважалися субпалліальнимі, є аналогами різних зон кори ссавців. Ймовірно, вони мають палліальное походження, і таким чином в мозку рептилій і птахів освіти палліум займають набагато більший обсяг, ніж вважалося раніше. Палліум рептилій представлений дорсальним вентрікулярную гребенем (DVR, dorsal ventricular ridge) з нідопалліумом (неостріатум по старій номенклатурі). У субпалліуме виділяють паллідум і стріа- тум, а також добре розвинену мигдалину, септальних ядра і прилегле ядро.
За сучасною номенклатурі палліум кінцевого мозку птахів включає гіперпалліум, мезопалліум, нідопалліум і аркопалліум (рис. 16.10).
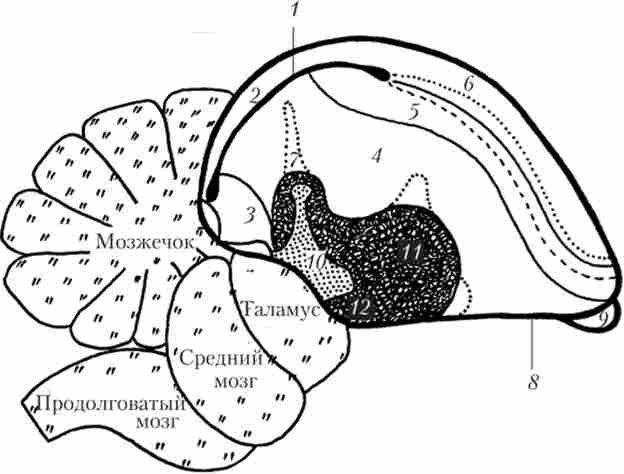
Мал. 16.10. Основні структури кінцевого мозку птахів (по Е. D. Jarvis1):
Гіперпалліум - структура, яка простежується тільки у птахів. Його ростральні відділи утворюють особливу дорсальне піднесення - Wulst. передній Wulst є областю соматосенсорного аналізу, задній Wulst пов'язаний з багатьма зоровими функціями, в тому числі з бінокулярний зір, просторової орієнтації, контролем за власними рухами щодо подій у зовнішньому середовищі (це аналоги соматосенсорної і зорової кори ссавців). У нідопалліуме знаходиться поле L - вищий центр аналізу складних звуків. Показано, що клітинне будова гіперпалліума, і особливо Wulst, корелює зі здібностями до навчання, пам'яті і елементарної розумової діяльності. Субпал- ліум птахів включає паллідум і стриатум, септальних область, прилегле ядро і ін.
Так як розумову діяльність тварин головним чином пов'язують з палліальнимі формаціями, сучасні уявлення про будову мозку птахів набагато краще узгоджуються з відомими фактами про їх високорозвинених психічних здібностях. Треба відзначити, що у самих «розумних» птахів (ворони, папуги, хижі) особливо добре представлені нідопалліум і мезопалліум.
Що стосується ссавців, то з будовою їх кінцевого мозку ми вже знайомі (див. Гл. 9). З точки зору філогенезу треба відзначити, що для цієї групи хребетних характерно надзвичайний розвиток неокор- текса, що супроводжується збільшенням таламуса і формуванням нових зв'язків з цією структурою. У новій корі з'являються пірамідні нейрони, які посилають свої волокна безпосередньо в СМ, в результаті чого формується пірамідна система, яка здійснює прямий контроль за рухами на відміну від многозвенной екстрапірамідної системи. Більш точна регуляція рухів забезпечується також і двосторонні зв'язки неокортексу з мозочком.
Серед різних ссавців спостерігаються відмінності в будові кінцевого мозку, і перш за все - кори великих півкуль. Ці відмінності пов'язані як з цитоархітектоніку кори, так і з топографією часткою кори, а також її борозен і звивин. У більшості ссавців в кожній півкулі виділяють тільки чотири частки - лобову, тім'яну, скроневу і потиличну; островковая частка з'являється тільки у приматів. Поділ півкуль на долі у багатьох ссавців носить відносний характер, так як борозни і звивини відрізняються великим як міжвидові, так і внутрішньовидових різноманітністю.
Виразність часткою у різних загонів також відрізняється. Вивчення цитоархітектоніки, а також аферентів і ефферентов коркових полів у різних груп призводить до висновку про великий невідповідність їх кірковим полях людини.
Найбільш загальна закономірність у відмінностях в будові кори пов'язана з кількістю борозен і звивин. За цим принципом кору ділять на гладку, практично не має борозен і звивин, і гіріфіцірованную (Gyrus - звивина) складчасту (див. рис. 16.9 - кролик і собака). Гладка кора характерна, наприклад, для комахоїдних, гризунів, Зайцеподібні, кажанів. У такій корі присутні тільки дві борозни - бічна і нюхова. Прикладами тварин з складчастої корою є копитні, хижаки, китоподібні і, звичайно, примати. Зрозуміло, що поява борозен і звивин збільшує корковую поверхню і, відповідно, здатності мозку до аналітико-синтетичної діяльності.
У зв'язку з цим виділяють три рівня організації інтеграційних систем мозку. Перший (у гризунів) характеризується відсутністю чіткої диференціації кори на специфічні проекційні і асоціативні зони, а таламуса - на проекційні і асоціативні ядра. Для другого рівня (у хижих) характерна поява в корі розвинених лобових і тім'яних асоціативних зон і відповідних структур таламуса - дорсоме- діальной ядра (DM), проектується в лобову кору, плодів та овочів LP і Pul. (Див. Параграф 8.1), що посилають волокна в тім'яну кору. У тварин третього рівня (примати) асоціативні зони розвинені максимально. Нагадаємо, що у людини вони займають більшу частину корковою поверхні.
На закінчення треба відзначити, що, незважаючи на загальну тенденцію в розвитку мозку, в кожному класі спостерігається значна варіабельність в його будові, пов'язана з морфофизиологічної організацією і способом життя тварини. У кожній групі хребетних є велика кількість способів організації мозкових структур, що ведуть до виконання подібних функціональних завдань.