| Головна |
| «« | ЗМІСТ | »» |
|---|
Крім виділення в корі великих півкуль часткою і звивин всю кору ділячи г на 52 поля відповідно до їх функцією і цитоархітектоніку, т. З. кількістю, величиною, формою і розподілом нейронів. Відрізняються поля і міелоархітектоніка, т. Е. Розподілом нервових волокон (рис. 9.9). Залежно від функцій, онтогенезу, особливостей цито- і міелоархітектоніка все коркові поля прийнято розділяти на первинні, або проекційні, вторинні і третинні, або асоціативні.
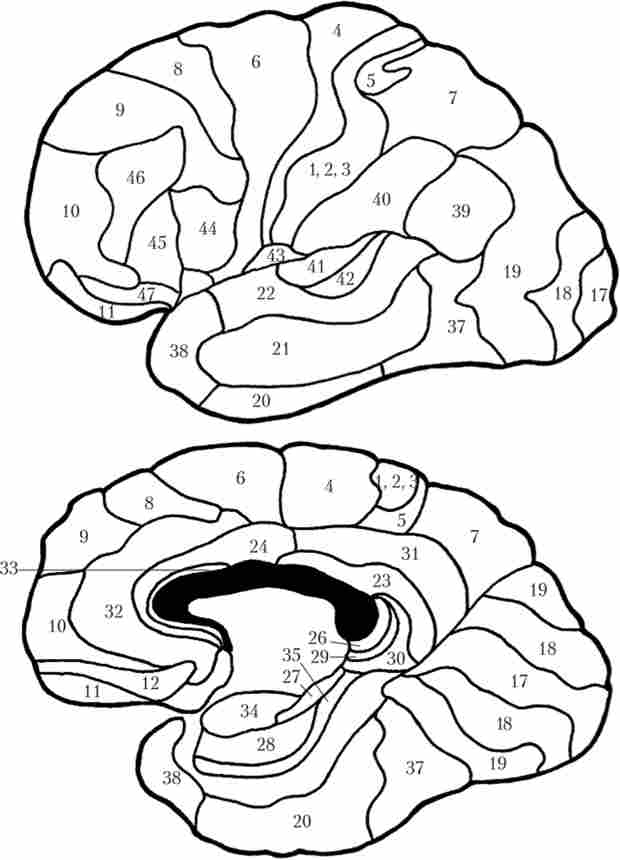
Мал. 9.9. Поля великих півкуль по Бродману
первинні поля отримують прямі входи від відповідних таламичних проекційних ядер, причому для цих нулів характерна тонична організація (див. параграф 11.3).
jвторинні поля оточують первинні і розглядаються як їх периферичні відділи. Крім прямих аферентів з таламуса вони отримують входи і від своїх проекційних зон.
Первинні і вторинні поля діляться на сенсорні і моторні.
первинна моторна (рухова) зона розташована в прецентральной звивині лобової частки кори великих півкуль попереду центральної борозни (поле 4); вторинна, або премоторная, зона (Поле 6) лежить безпосередньо перед первинної. Вони отримують входи від рухових проекційних ядер таламуса. Більшість тих, які сходять шляхів кори великих півкуль виходить з поля 4. В основному ці шляхи починаються у внутрішньому пирамидном шарі, який виражений тут значно сильніше, ніж в інших зонах (агранулярна кора). Для поля 4 характерна соматотопія (див. Параграф 5.4), т. Е. На ньому можна намалювати «карту тіла» - сусідні ділянки первинної моторної кори відповідають за рух сусідніх м'язів тіла. Таку соматотопичну карту прийнято називати «гомункулус» (чоловічок). Карта відрізняється певними спотвореннями пропорцій людського тіла (рис. 9.10). Пов'язано це з тим, що деякі м'язи (пальців, мімічні) повинні виконувати набагато більш тонкі руху, тому для управління ними необхідна більша кількість нейронів. Премоторная кора передає сигнали до первинної моторної кори, запускаючи рухові команди, що йдуть але спадним трактах.
До сенсорним зонам кори відносять нуля, в які приходять волокна від проекційних сенсорних ядер таламуса. Це зони коркового представництва сенсорних систем. Для цих зон характерні дуже сильний розвиток IV шару кори, куди приходять таламичні афференти, і в той же час погано виражений V шар (гранулярна кора).
Для кожної сенсорної системи існують свої проекційні зони. зорова зона знаходиться в потиличній частці кори великих півкуль. Вона розташована на ділянці під назвою «клин» на медіальної поверхні. Первинна зорова кора займає поле 17, вторинна - поля 18 і 19. Вони отримують афференти від латерального колінчастого тіла таламуса. слухова зона розташована у верхній скроневій звивині. Первинна (в поле 41) і вторинна (в поле 42) зони отримують входи від медіального колінчастого тіла. смакові зони знайдені в поле 43 (нижня частина постцентральна звивини) і в инсулярной корі.
Великий ділянку займає зона шкірно-м'язової чутливості (iсоматосенсорная кора). Вона розташована позаду центральної борозни, в постцентральна звивині тім'яної частки кори (поля 1 і 3 - первинна кора, поля 2 і 43 - вторинна кора). Сюди приходять афференти від вентробазальіого комплексу таламуса. Так само як і моторна кора, шкірно-м'язові проекції організовані по соматотопичної принципом. І так само як і моторний гомункулус, сенсорний чоловічок має спотворені пропорції. Справа в тому, що кількість нейронів, які отримують інформацію від певної ділянки тіла, прямо пропорційно щільності рецепторів на цій ділянці. Щільність же рецепторів залежить від значущості інформації, одержуваної від тієї чи іншої ділянки шкірної поверхні. Тому на «карті тіла» в корі відзначаються непропорційно великі зони кистей рук і губ, але дуже маленькі зони спини, живота і т. Д. (Рис. 9.10).
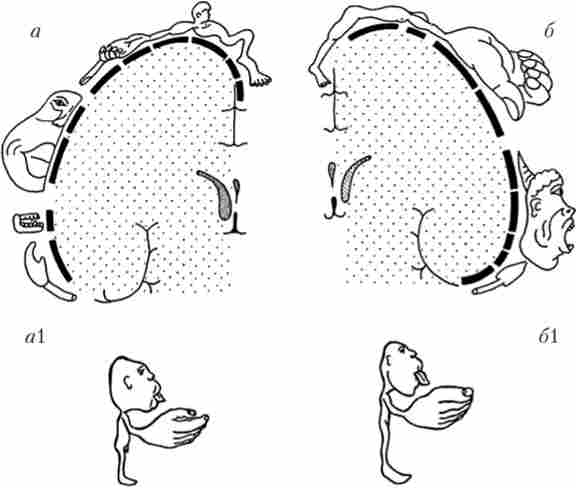
Мал. 9.10. Представництво зон шкірної чутливості і рухових зон в корі великих півкуль:
а - проекційна зона шкірної чутливості; 6 - моторна проекційна зона. Невідповідність пропорцій проілюстрована у вигляді сенсорного (а) І моторного
(61) гомункулуса (чоловічка)
До третинної (асоціативної) корі віднесені області, яким можна приписати будь-яких переважно сенсорних або рухових функцій. Вони отримують афференти від асоціативних ядер таламуса, а також від первинних і вторинних полів. Площа асоціативних зон безпосередньо пов'язана з рівнем вищої нервової діяльності. У людини асоціативні зони займають більше половини всієї поверхні кори. Ці зони пов'язують (асоціюють) один з одним сенсорні і рухові зони і одночасно служать субстратом вищих психічних функцій.
Основні неспецифічні області кори великих півкуль це тім'яно-скронево-потилична, або зона ТРО (Temporalis - скронева, parientalis - тім'яна, occipitalis - потилична), яка знаходиться на кордоні цих часток; префронтальная (лобова частка кори, за винятком полів 4 і 6); лімбічна (склепінчаста звивина) асоціативні зони. Якщо спрощено уявити їх функції, кожна з цих областей дуже багато важить відповідно для наступних інтеграційних процесів: вищих сенсорних функцій і мови; вищих рухових функцій і запуску поведінкових актів; пам'яті і емоційного поведінки.
Найбільші асоціативні поля належать лобної долі. Нагадаємо, що в числі інших вона отримує афференти від ядра таламуса MD. Па її нижній медіальної поверхні знаходиться орбито-фронтальна, або орбітальна, кора (Поля 11 і 12), яка вважається важливою частиною лімбічної системи. На латеральній поверхні розташована префронтальная асоціативна область, або префронтальная кора (Поля 8-10, 44-47), функції якої вельми багато. Наприклад, поля 44-46 пов'язані з регулюванням речедвігательних функцій, нейрони поля 8, яке іноді відносять до вторинної рухової корі, пов'язують з фокусуванням зорової уваги. Поразка префронтальної кори приводить до нездатності планувати свої дії, апатії, безініціативності.
Зона ТРО отримує афференти від асоціативних ядер таламуса LP і LD. У цю зону приходять сигнали від різних сенсорних зон кори, в результаті чого там формується полімодальний сенсорний образ. При порушенні роботи тім'яно-скроневої зони можливий розвиток різноманітних агнозий (порушень сприйняття). У цій же зоні є поля, пов'язані зі сприйняттям мови (див. Нижче).
Класифікація полів на первинні, вторинні і третинні не є універсальною і викликає ряд питань. Наприклад, нюхові зони кори але функціональним критеріям можна розділити на первинні, вторинні і третинні, але будова палео-і архікортікальних утворень, де знаходяться ці зони, принципово відрізняється від будови неокортексу. Або поле 43 розглядається як первинна смакова кора і в той же час як вторинна сснсомоторная зона. Крім того, в неокор- текс є зони, що мають проміжні риси як в анатомічному, так і у функціональному відношенні. Проте розподіл кіркових полів на первинні, вторинні і третинні зберігає свою актуальність і широко використовується в анатомо-фізіологічної літературі.
В даний час все більшого значення набуває модульний принцип організації кори великих півкуль.
Модуль - це вертикально орієнтована група нейронів, що виконують певну функцію, що відрізняється від функцій сусідніх модулів.
Нейрони кожного модуля об'єднані пучком апікальних дендрі- тов (від 2-3 до 20) пірамідних клітин. Між апікальними дендрі- тами і тілами цих пірамід існують численні електричні синапси, завдяки чому координується робота цих клітин. Внутрімо- дульні зв'язку здійснюються різними непіраміднимі нейронами, що входять до складу модуля. Нейрони модуля але своїми функціями діляться на три групи: аферентні нейрони, на яких утворюють синапси волокна, що приходять з внемодульних утворень; еферентні нейрони, аксони яких виходять з модуля; інтернейрони, що реалізують Внутрімодульное зв'язку. Прикладом модулів можуть служити колонки в зоровій корі (див. Параграф 13.4).
Вперше модулі були описані для кори великих півкуль. Надалі вони були описані в різних відділах ЦНС багатьох хребетних, де вони можуть приймати іншу форму, але складатися з таких же трьох груп нейронів. Це дозволяє припускати, що модуль є структурно-функціональною одиницею будь-якого нервового центру.
Хоча за своєю будовою праве і ліве півкулі людини відрізняються незначно, для них характерна функціональна асиметрія, т. е. вони виконують різні функції. В першу чергу це відноситься до асоціативних зон кори. У повсякденному житті ці відмінності не помітні, тому що інформація легко переходить з півкулі в півкулю через коміссури мозку (в першу чергу, через мозолисте тіло). Уявлення про відмінності у функціях півкуль склалися при спостереженнях за хворими з односторонніми ураженнями мозку і в спеціальних експериментах, де інформація надходила тільки в одне з півкуль.
Виявилося, що ліва півкуля (по крайней мере, у правшів) в більшій мірі пов'язане з промовою, абстрактно-понятійним мисленням, математичними здібностями. Права півкуля переважно керує образним мисленням і в значній мірі визначає такі властивості, як музикальність, розпізнавання складних зорових образів, вираз і сприйняття емоцій. При ураженні деяких областей правої півкулі відзначаються глибокі порушення орієнтації в просторі, наприклад, хворі не можуть знайти дорогу в будинку, в якому прожили багато років. Деякі пацієнти з ушкодженнями правої півкулі не можуть впізнавати знайомі обличчя, а іноді взагалі не можуть впізнавати людей.
У лобової і скроневої областях лівої півкулі у правшів знаходяться центри мови. У верхньої скроневої звивини на кордоні зі слуховий корою знаходиться зона Верніке (Поле 22). Це зона сприйняття почутої мови. При ураженні цієї зони людина перестає розуміти чуже мовлення, але розуміє мову письмову або жестовую. У префронтальної корі (поля 44, 45) розташована зона Брока. Це руховий центр мови. При його ураженні хворий чує і розуміє мову, але сам говорити не може. При цьому хворий не втрачає здатності до письмової та жестової мови. Ці дві зони тісно пов'язані між собою волокнами, що проходять в складі крючковидного пучка переднього мозку. Тому при ураженні зони Верніке нерідко спостерігаються і проблеми з підбором і проголошенням потрібних слів.
На закінчення необхідно підкреслити, що мозок має надзвичайно великі компенсаторні можливості. Звичайно, багатьом його зонам (ядер) притаманні врожденно обумовлені функції (особливо це характерно для первинних сенсорних зон, центрів довгастого мозку і т. П.). Однак дуже багато областей, в першу чергу коркові освіти, набувають конкретні «обов'язки» у міру дозрівання і навчання ЦНС. Це властивість мозку зумовлює можливість того, що при ураженні різних ділянок існує можливість, що інші відділи ЦНС візьмуть на себе виконання відповідних функцій.