| Головна |
| «« | ЗМІСТ |
|---|
Раніше, в Середні століття, існувало уявлення, що психічні функції локалізовані в трьох мозкових шлуночках. І тільки в 1822 р німецький анатом Ф. Галль сформулював інший підхід до цієї проблеми в рамках науки френології, засновником якої його вважають (див. Гл. 1). Було висунуто припущення, що за психічні функції відповідає кора б. п. Однак ця прогресивна думка привела до ряду помилок, в результаті яких френологія не мала успіху вже в той час. Згідно з підходом з позицій френології, форма черепа відображає будову лежить під ним мозкової тканини. Галль вважав, що за формою черепа (горбки, шишечки, вм'ятини і т. П.) Можна судити про ступінь розвиненості тієї чи іншої зони кори і, отже, про розвиток тієї чи іншої психічної функції.
Великий внесок у розвиток уявлень про локалізацію функцій в корі б. п. внесли канадські вчені В. Пенфилд і Г. Джаспер. У 30-х рр. XX ст. вони з метою лікування епілепсії розробили спеціальну операцію по видаленню епілептичного вогнища, але для більш точної операції вони повинні були бути впевнені, що ні видалять «зайвих» ділянок кори. Для уточнення функцій різних областей кори вони проводили пряме електричне роздратування головного мозку слабким струмом. В результаті експериментів Пенфилда і Джаспера було складено досить докладні карти кори, які використовуються досі.
З функціональної точки зору нову кору прийнято поділяти на три типи зон (областей): проекційні (сенсорні), асоціативні, рухові (моторні).
Проекційні зони кори здійснюють вищий рівень аналізу зовнішніх і внутрішніх сенсорних сигналів. Вони отримують інформацію безпосередньо від специфічних (релейних) ядер таламуса. Кожна сенсорна зона має топичний принцип організації - кожній ділянці рецепторной поверхні відповідає чітко певну ділянку в проекційної корі. У кожній сенсорної зоні зазвичай виділяють первинні, вторинні і в деяких випадках навіть третинні проекційні зони. Основними сенсорними зонами є зорові, слухові і соматосенсорні області.
Зорові проекційні зони займають потиличну область кори (поля 17, 18 і 19 по Бродману). На рівні первинної зорової зони відбувається детектування окремих ознак зорового образу. Вторинні і третинні зорові зони здійснюють взаємодію зорового сигналу з інформацією від інших сенсорних систем (слуховий, соматосенсорної і т. Д.).
Слухові проекційні зони знаходяться в скроневих областях кори. Первинні слухові зони займають поля 41 і 42, а вторинні - поля 21, 22 і 52. У первинних слухових областях відбувається аналіз звуків різної частоти, причому всі нейрони одній вертикальній колонки реагують на звук однієї певної частоти. Тут також відбувається аналіз тони, гучності і характеру коротких (до 100 мс) звуків. У вторинних слухових зонах здійснюється оцінка значимості звукових сигналів (видова і індивідуальна). Особливе значення для людини має область скроневої кори лівої півкулі (поле 22), яка має назву зона Верніке. Ця область відповідає за розуміння і сприйняття мови (своєї та чужої) і є сенсорним центром мови. При пошкодженні цієї області у людини спостерігається так звана сенсорна афазія, або афазія Верніке, при якій людина говорить багато, але безглуздо, так як погано розуміє як чужу, так і свою промову.
Соматосенсорні проекційні області здійснюють вищий рівень аналізу шкірної, м'язової і вісцеральної чутливості. Первинні соматосенсорні області розташовані в області центральної борозни (нуля 1, 2 і 3 але Бродману, задня центральна звивина). Ця область кори має чітку топичну організацію - кожна ділянка тіла людини представлений своєю ділянкою в соматосенсорной корі (рис. 15.4).
Вторинна соматосенсорная зона невелика, розташовується в латеральній борозні. Туди надходять сигнали і від вісцеральних рецепторів (від внутрішніх органів).
Асоціативні зони кори не мають спеціалізованих входів, не мають конкретних проявів їх стимуляції або руйнування, тому їх іноді називають «мовчати» зонами. При цьому саме асоціативні зони кори в ході філогенезу набувають все більшого значення в регуляції складних форм поведінки і у приматів займають значну частину неокортексу. Основними асоціативними зонами є тім'яна кора (поля 5, 7, 39, 40 по Бродману) і лобова кора (поля з 8-го по 12-е).
Тім'яна асоціативна зона забезпечує відтворення цілісних образів предметів і явищ, здійснює інтеграцію аферентних потоків від різних сенсорних систем. На нейронах тім'яної зони відбувається конвергенція аферентних потоків різної сенсорної модальності, що створює оптимальні можливості для сприйняття цілісного образу предмета і його просторово-часових відносин з іншими предметами. Більшість нейронів тім'яної зони реагують на стимули двох або трьох модальностей. Тім'яна кора отримує сигнали від проекційних областей кори по асоціативним волокнах і від асоціативних ядер таламуса (від подушки і вентробазального комплексу). Більше число еферентних виходів з тім'яної області йде в моторну кору, де і відбувається формування команди довільного руху.
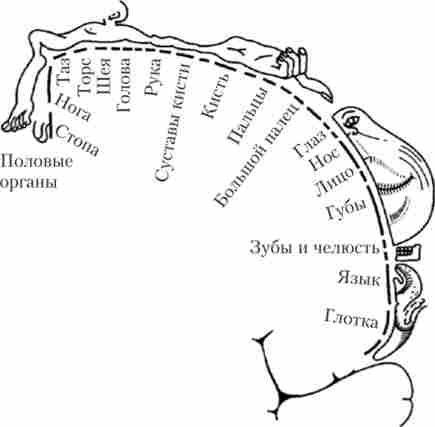
Мал. 15.4. «Сенсорний гомункулюс» - проекції частин тіла в соматосенсорной корі в області постцентральной звивини
Особливо сильно виражена інтегративна функція тім'яної кори у людини. При пошкодженні цієї області порушується здатність комплексного сприйняття предметів у всій сукупності їх ознак, порушується диференціювання предметів, їх просторове розрізнення. В першу чергу порушується здатність синтезувати окремі компоненти в складну систему цілеспрямованої поведінки.
Стимуляція ділянок тім'яної кори може привести до змін в процесах пам'яті (можна викликати спогади про події, в яких людина давно брав участь), причому спогади супроводжуються відповідними емоційними переживаннями.
Лобові асоціативні зони сформовані повністю тільки у приматів і людини. Нейрони цих зон полімодальних і мають безліч зв'язків з іншими корковими областями, а також з підкірковими структурами. У людини передні ділянки лобових часток беруть участь в реалізації найбільш складних процесів, пов'язаних з безпекою особистості, формуванням соціальних відносин, адекватністю поведінки. Ділянки лобової кори пов'язані з організацією цілеспрямованого поведінки: програмування, прогнозування наслідків і контроль поведінки, а також з функцією прийняття рішення.
Лобові частки кори б. п. у людини беруть участь в організації мовленнєвої діяльності - нижня третина задньої лобової звивини містить моторну мовну зону (Зону Брока), відповідальну за проголошення слів. При ураженні зони Брока настає моторна афазія, при якій людина з працею вимовляє слова, але розуміння мови не порушено. При пошкодженні інших сусідніх ділянок можуть спостерігатися порушення здатності писати (аграфия) і здатності читати вголос.
Пошкодження і лобових, і тім'яних зон кори б. п. позначаються на найбільш складних формах діяльності мозку, однак, до схожих порушень можуть привести і пошкодження інших структур мозку, тому прийнято вважати, що психічна діяльність не локалізована в якихось окремих структурах, а є функцією всього мозку.
Рухові (моторні) зони розташовані в прецентральной області (поля 4 і 6 по Бродману), а також на медіальної поверхні є ще додаткова моторна область. Електрична стимуляція різних ділянок моторної кори приводить до скорочень окремих м'язів протилежного боку тіла. Ці області кори мають строгу топичну організацію - кожній ділянці кори відповідає своя група м'язів ( «моторний гомункулюс», рис. 15.5).
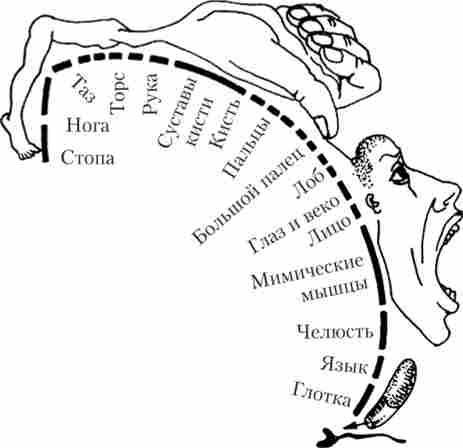
Мал. 15.5. «Моторний гомункулюс» - представництво м'язів різних частин тіла в моторній корі в області прецентральной звивини
Нейронні угруповання моторної кори, пов'язані з рухами різних м'язів, займають різні за розміром площі і розподілені нерівномірно. Великі ділянки кори відповідають за рухи пальців рук, кистей, мови, м'язів обличчя і істотно менше за руху великих м'язів спини і нижніх кінцівок. Такий нерівномірний розподіл пов'язано з тим, що аксони пірамідних нейронів моторної кори дають найбільше число синаптичних контактів на мотонейрони спинного мозку, що відповідають за тонку і найбільш точну регуляцію чітких рухів кистей рук, обличчя, язика.
Пошкодження моторної кори у людини призводить до паралічів відповідної мускулатури. При односторонніх ураженнях відбувається поступове відновлення рухових функцій за рахунок компенсаторних процесів в протилежному півкулі, при двосторонньому пошкодженні відновлення не відбувається.
Аферентні (вхідні) сигнали в моторну кору приходять з сенсорних зон обох півкуль, від асоціативних областей кори і з підкіркових структур. Найбільш чітко організовані проекції в моторну кору з вентролатерального ядра таламуса. Велике значення має надходить в моторну кору інформація від м'язів, суглобів і сухожиль (пропріорецепція).
Нейрони моторної кори згруповані у вертикальні колонки, які керують роботою невеликих груп м'язових волокон. Існують окремі колонки моторної кори, пов'язані з швидкими (фізичними) і повільними (тонічними) рухами. Еферентних виходами таких колонок служать відростки гігантських пірамідних нейронів 5-го шару кори. Їх аксони формують пірамідні тракти, що закінчуються на мотонейронах різного рівня спинного мозку протилежного боку (пірамідна система контролю рухів). Ці тракти беруть участь в здійсненні довільних рухів.
У моторній корі починається також екстрапірамідна система. Аксони цієї системи з кори направляються до базальних гангліїв, мозочка, червоного ядра середнього мозку, ядер ретикулярної формації і іншим структурам стовбура мозку.
У той же час кора б. п., функціонуючи як єдине ціле, здійснює процеси обробки чутливої (сенсорної) інформації, формує рухові команди. Кора забезпечує індивідуальне пристосування людини до мінливих умов внутрішнього і навколишнього середовища. Завдяки гнучкій пристосовності до постійно мінливих обстановці організм людини зберігає життєздатність і життєдіяльність.