| Головна |
| «« | ЗМІСТ | »» |
|---|
Генетичні системи багатоклітинних тварин засновані переважно на раздельнополости і перехресному заплідненні, за винятком випадків гермафродитизму, при якому жіночі та чоловічі статеві органи утворюються у одній і тій же особини. Гермафродит поширений серед безхребетних: кишковопорожнинних, плоских хробаків, деяких молюсків. У хребетних це деякі риби, наприклад морський окунь. При гермафродитизме в зв'язку з різночасністю дозрівання яйцеклітин і сперматозоїдів однієї і тієї ж особи як правило відбувається перехресне запліднення.
Вищі тварини також потенційно бісексуальні, однак хромосомний механізм визначення статі обумовлює диференціацію чоловічих або жіночих особливостей організму. Розрізняють первинні статеві ознаки - будова гонад, а також ті фізіологічні і морфологічні особливості, які визначають нормальний розвиток гамет і їхнє злиття при заплідненні, і вторинні статеві ознаки, характерні для даної статі, але безпосередньо не пов'язані з процесом статевого розмноження.
Зачатки гонад у ембріонів тварин містять два типи тканин - кортекс (Зовнішній шар) і медулла (Внутрішній шар). В процесі діффе- ренціровкі, згідно хромосомної детермінації статі, у жіночих особин кортекс розвивається в яєчники, а у чоловічих особин з медулли утворюються насінники. Потенційну бісексуальність тварин наочно демонструють експерименти по перевизначення статі в онтогенезі. Кастрація півня приводить до розвитку у нього ознак курки. Якщо такий курці знову пересадити насінники, то відновлюються ознаки чоловічої статі.
У великої рогатої худоби відомий феномен фрімартінізм. При народженні різностатевої двійні у корови в деяких випадках внутрішні органи телиці набувають ознак чоловічої статі, хоча зовнішні генІталії влаштовані за жіночим типом. Такі фрімартінамі завжди стерильні. Їх поява пояснюється тим, що чоловічі гормони бичка, що розвивається нормально, починають виділятися раніше, ніж жіночі гормони телиці, і скасовують підлогу жіночого ембріона.
Штучне гормональне перевизначення статі може бути тимчасовим, як, наприклад, у курей при обробці яєць жіночим гормоном - естрогеном, або постійним, як це показано для деяких земноводних і риб. Так, у акваріумний рибки Otyzias latipes дією жіночого статевого гормону на новонароджених малюків вдалося повністю перевизначити підлогу. В результаті генетичні самці (АТ) функціонували як самки і приносили потомство з розщепленням по генам, зчепленим з підлогою, якого слід було б очікувати при схрещуванні двох самців. У даного виду риб особини YY життєздатні.
Початкові стадії формування попередників чоловічих і жіночих статевих клітин - гоніальних клітин - подібні. Схема сперматогенезу і оогенезу представлена на рис. 7.3.
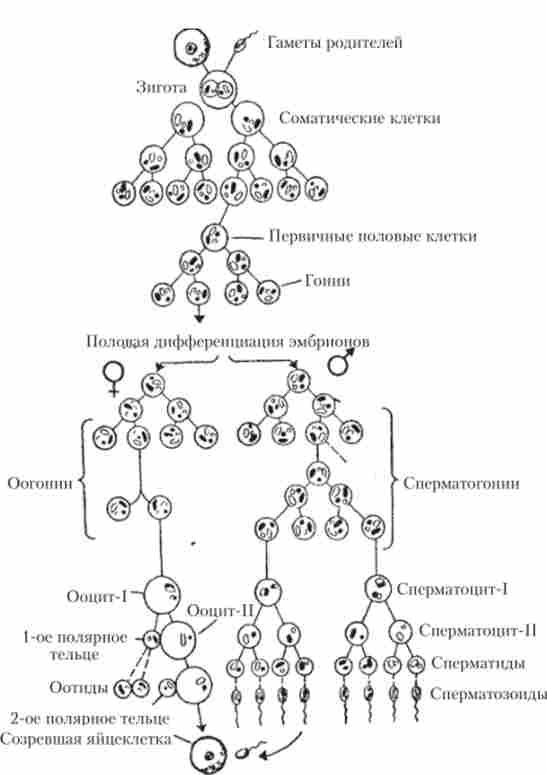
Мал. 7.3. Порівняльна схема розвитку чоловічих (сперматогенез) і жіночих (оогенез) статевих клітин
У сім'яниках сперматогонии після періоду зростання перетворюються в сперматоціти I порядку, які зазнають I поділ мейозу, і в результаті утворюються сперматоціти II порядку. У них проходить 11 поділ мейозу, в результаті чого виникають сперматіди. Так кожен диплоїдний сперматоцит дає чотири гаплоїдні клітини. Далі сперматіди в результаті процесу сперміогенеза перетворюються на сперматозоїди. При цьому утворюється головка сперматозоїда, представлена майже виключно гаплоидним ядром, а всі елементи цитоплазми беруть участь у формуванні хвоста і апарату, що забезпечує рухливість сперматозоїда.
Розвиток жіночих статевих клітин - яйцеклітин - відбувається в яєчниках. Як яйцеклітини, так і сперматозоїди зазнають два ділення мейозу. Істотною відмінністю при цьому є те, що ооцити I по- рядка, в які перетворюються оогонии в результаті більш тривалого зростання, утворюють при I поділі мейозу дві гаплоїдні клітини з неоднаковим кількістю цитоплазми: одну велику клітку - ооцит II порядку і невелике перших полярних тільце. При II поділі мейозу кожен ооцит II утворює одну оотіду і другий полярне тільце. Перше полярне тільце також ділиться. В результаті мейозу ооцит I утворює чотири оотіди, але тільки одна з них, дозріваючи, перетворюється в яйцеклітину. Три залишилися клітини абортивного.
Надалі при заплідненні в яйцеклітину проникає тільки ядро сперматозоїда - чоловічої пронуклеус. Він зливається з жіночим пронуклеусом - ядром яйцеклітини. Сперматозоїд може проникати в яйцеклітину не тільки на стадії зрілості, але у деяких тварин - і на різних стадіях формування ооцита I і ооцита II. Проте при заплідненні зливаються тільки гаплоїдні ядра, що завершили мейоз.
Надалі зигота розвивається тільки за рахунок цитоплазми яйця, а сперматозоїд вносить в зиготу тільки свою половину хромосомного набору.
Розглянутий тип запліднення носить назву анізогамії, або гетерогамії. При цьому нерівність зливаються гамет стосується тільки цитоплазми, в той час як ядерний матеріал посту Пает від батька і матері. Нерівність гамет по цитоплазмі дозволяє досліджувати відносну роль ядра і цитоплазми в детермінації ознак. Для цього використовують реципрокні схрещування. Подальше перерозподіл батьківського і материнського матеріалу відбувається в мейозі, проте можливі випадки митотичної рекомбінації, які можуть мати генетичне значення тільки у організмів, що розмножуються вегетативним шляхом.
Таким чином, життєвий цикл багатоклітинних тварин представлений диплоидной стадією (див. Рис. 7.1), а гаплоидни тільки статеві клітини. Виняток становлять тварини з гапло-диплоїдним життєвим циклом.