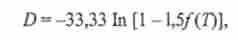
| Головна |
| «« | ЗМІСТ | »» |
|---|
Першими мікроорганізмами, які почала освоювати генетика в 1930-і рр., Були гриби, насамперед Neurospora, і різні види дріжджів роду Saccharomyces.
Мікроорганізми як правило не відрізняються різноманітністю морфологічних ознак, але це здається незручність компенсує можливість наблизитися до генетичного контролю метаболізму клітини за допомогою отримання мутантів, ауксотрофності (недостатніх) по різним органічних сполук, або мутантів, здатних засвоювати екзогенні джерела вуглецю і азоту. Тим самим концепція елементарного ознаки, або фена, конкретизується у вигляді однієї біохімічної реакції і далі - у вигляді активності ферменту.
Метод селективних середовищ, широко використовуваний в мікробіології, дозволяє реєструвати рідкісні події: мутації і рекомбінації, - і створювати умови, при яких виростають тільки цікавлять експериментатора варіанти. Селективні середовища широко використовують для відбору гібридів між гаплоїдними батьками, маркованими комплементарними рецесивними мутаціями ауксотрофності.
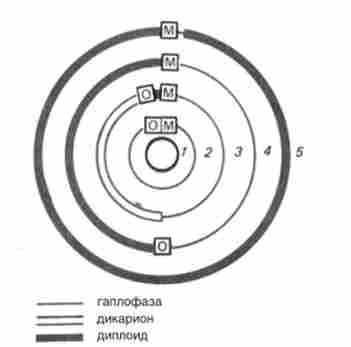
Нижче представлені п'ять основних варіантів життєвих циклів грибів (рис. 7.7). Серед грибів з статевим розмноженням спостерігаються варіації від повністю гаплоидного циклу (рис. 7.7, У, 2), як, наприклад, у Neurospora, до повністю диплоидного (рис. 7.7, 5), як у гомоталлічних штамів Saccharomyces cerevisiae (Рис. 7.8).
Для більшості міцеліальних грибів характерна наявність декількох ядер в загальній цитоплазмі. Ці ядра можуть мати різні генотипи. У такому випадку за аналогією з гетерозиготой говорять про гетерокаріо- нах (Рис. 7.7, 5).
У загальному вигляді мітоз і мейоз у грибів відбуваються так само, як у вищих еукаріот, але без руйнування ядерної мембрани і, мабуть, без щільної компактизации хромосом, оскільки останні зазвичай невиразні в світловому мікроскопі.
Мал. 7.7. Основні типи життєвих циклів у грибів в схематичному поданні: 1 - асексуальний; 2 гаплоїдний; 3 - гаплоїдний з гстсрокаріотічної стадією;
4 - гапло-диплоїдний; 5 - диплоїдний; М - мейоз; Про - запліднення
Мал. 7.8. Життєвий цикл дріжджів Saccharomyces cerevisiae: а - гетероталлічних, б - гомоталлічних
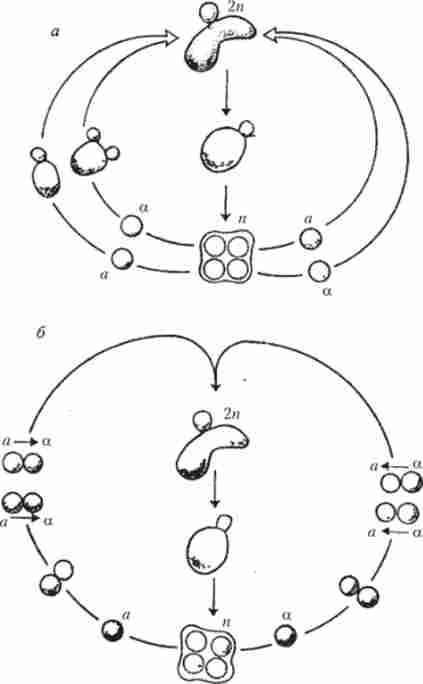
Несумісність у грибів. Залежно від способу генетичної детермінації несумісність у грибів буває гомогенної і гетерогенної. У першому випадку можливі схрещування між різними клітинами (міцелій), а в другому - між однаковими, по генам несумісності. Гомогенна несумісність, в свою чергу, підрозділяється на біполярну і тетраполярная (Рис. 7.9).
Мал. 7.9. Класифікація типів несумісності у грибів
Типовий представник грибів з гомогенної біполярної несумісністю - дріжджі Sacch. cerevisae (Рис. 7.8), у яких цітогамія і подальша кариогамия відбуваються тільки між 'клітинами (або аскоспорами) протилежних типів спарювання а і а, детермініруемих алелями локусу. До цього ж типу несумісності належить і Neurospora crassa.
Тетраполярная гомогенна несумісність характерна для базі- діоміцетов, зокрема для Shizophyllum соттіпае. При цьому типі несумісності нормальний статевий процес відбувається тільки між штамами, неоднаковими відразу за двома чинниками несумісності: А і В. При розходженні тільки по одному з факторів Л =% В * або А*, В = (Де = означає однакові, * - розрізняються фактори) гіфи зливаються і здійснюється лише частина реакцій статевого морфогенезу, що не завершується злиттям ядер. Гомогенний тип несумісності у грибів схожий з гомо- морфічного типом несумісності вищих рослин.
гетерогенна несумісність означає, що статевий процес відбувається тільки між штамами, однаковими за такими чинниками несумісності. При цьому несумісність контролюється як алелями одного гена, так і різними генами. Цей складний контроль може накладатися на гомогенну несумісність, як це спостерігається у Podospora anserina. У межах кожної географічної раси цього гриба відзначено два типи спаровування: «+» і «-». Нормальний статевий процес завершується тільки тоді, коли схрещуються штами «+» і «-» типів спарювання однакові по аллелям гена t. У Р. anserina існує і неалельних механізм гетерогенної несумісності, менш досліджений, ніж алельних. При неалельні механізмі несумісності для нормального схрещування необхідно, щоб штами були однаковими відразу за кількома генами (не менше чотирьох).
Різноманітність життєвих циклів і типів несумісності у грибів накладає відбиток і на прийоми, які використовуються при їх гибридологічний аналізі. У одних грибів статевий процес здійснюється на основі гетерогамії, як у нейроспори, що дозволяє ставити реципрокні схрещування; у інших - на основі ізогамії, як у дріжджів сахароміцетів. Поряд з статевим розмноженням існує повний або неповний пара- сексуальний цикл в залежності від виду грибів. Парасексуальний цикл - це процес об'єднання і подальшої рекомбінації генів на основі подій, що відбуваються в мітозі, а не в мейозі, без участі запліднення статевим шляхом.
Розглянемо два підходи, які зробили істотний внесок у розробку проблем загальної генетики: тетрадном аналізі і генетичному аналізі на основі парасексуальними процесу.
Зошитовий аналіз зіграв вирішальну роль в доведенні правила чистоти гамет, у вивченні кросинговеру на стадії чотирьох хроматид і в рішенні інших проблем.
На відміну від генетичного аналізу, заснованого на випадковою вибіркою продуктів розщеплення (у вищих рослин, тварин і більшості мікроорганізмів), де одиницею варіювання (вимірювання) служить особина або клітка, в тетрадном аналізі такою одиницею є тетрада, точніше, саме розщеплення в тетраде. При моногібрідномсхрещуванні типове розщеплення в тетрадах - співвідношення 2А : 2а, якщо в об'єкта четирехспоровие аски, як у дріжджів або Neurospora tetrasperma, а також тетради мікроспор, як у вищих рослин. У разі восьміспорових ас- ков (октад) це співвідношення перетворюється в 4А : 4а, як у N. crassa або Sordaria fimicola.
Розглянемо тетради впорядковані, як у N. crassa, і невпорядковані, як у Saccharomyces, де зазвичай неможливо встановити порядок аскоспор щодо направлення веретена двох мейотічних поділів. В обох випадках (для упорядкованих і неупорядкованих тетрад) при ди- гібридному схрещуванні, наприклад АВ х ab, в загальному вигляді розглядають три типи зошити: батьківський дітіп (Р), неродітельскій дітіп (N) і тетратіп (Г):
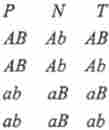
нехай гени А і В знаходяться в різних хромосомах. тетради Р і N можуть з'явитися, якщо в першому поділі мейозу збігається розщеплення
(Редукція) по Центромера і по обом факторам A / а і В / b. Ці випадки носять назву розщеплення при першому поділі мейозу. тетради Т з'являються тільки в результаті кросинговеру на ділянці ген A / а (І / або В / Ь) - центромера (рис. 7.10). На цьому факті засновано картування генів стосовно Центромера.

Мал. 7.10. Освіта тетрад Р, N і Т в залежності від наявності або відсутності кросинговеру між генами і відповідними центромерами. М, Ml - перше і друге ділення мейозу
якщо гени А і В не зчеплені між собою і вільно рекомбинируют зі своїми центромерами, т. е. досить віддалені від них, то легко переконатися, що співвідношення тетрад Р: N: Т - 1: 1: 4, оскільки тетради Т виникають в результаті чотирьох варіантів обміну на стадії чотирьох хро- матіди на ділянці ген-центромера.
Якщо обидва гена - А і В - зчеплені досить тісно зі своїми центромерами, то розщеплення при другому розподілі утруднено (кросинговер рідкісний), що призводить до зменшення частоти тетрад Т. тоді при Р = N, 2
7 < - можна визначити відстань генів від своїх центромер по частоті розщеплення при другому розподілі. Реально для неупорядкованих зошити відстань ген-центромера визначають при трігібрідного схрещуванні, при якому всі три гена виявляють зчеплення з різними центромерами. У разі упорядкованих зошити яку можна визначити і при моногібрідномсхрещуванні по розташуванню суперечка в лінійному аске.
якщо гени А і В зчеплені між собою, то тетради Р wN з'являються 2
ні з однаковою частотою: Р > N, Т -. У цьому випадку відстань між генами можна визначити також виходячи з частот Т.
У загальному вигляді формула для визначення відстані (D) між генами:
де / (7) - частота тетратіпов (похідна від функції Холдейна).
Відстань між генами і центромерами визначають при трігібрідного схрещуванні, користуючись тією ж формулою, однак тепер величина D - це сума відстаней двох генів від їх центромер. Гак визначають DABiDAC і Dbcj далі - відстань кожного гена від своєї центромери:
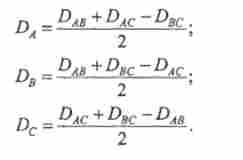
Відстань виражається в умовних одиницях - Стрейна. Найменування «Стрейн» походить від загального скорочення імен трьох генетиків, які зробили видатний внесок у вивчення рекомбінації: А. Стертеванта, А. Троу і Дж. Холдейна.
При гибридологічний аналізі у грибів можна використовувати випадкову вибірку гаплоїдних сегрегантов при звільненні аскоспор з асков. Конкретна тактика використання прийомів генетичного аналізу диктується його завданнями. Так, відстань між генами простіше визначити в випадковою вибіркою аскоспор, а зчеплення генів з центромерами - в тетрадном аналізі.
Генетичний аналіз при парасексуалиюм процесі. У багатьох грибів можливо злиття вегетативних гіф, в результаті чого гаплоїдні ядра штамів виявляються в загальній цитоплазмі. При генетичні відмінності між такими ядрами виникають гетерокаріони, які можуть тривалий час існувати, як, наприклад, у N. crassa. Цей прийом широко використовується для вивчення взаємодії між генами і алелями, між генами ядра і цитоплазмою. При цьому між алелями одного гена можуть встановлюватися рецессивно-домінантні відносини, в загальних рисах аналогічні відносинам в гетерозиготі. Однак в тих випадках, де це можливо порівнювати, гетерозиготи і гетсрокаріони іноді показують різний характер алельних взаємодій, що, мабуть, пов'язано мінімум з двома обставинами:
У деяких грибів, наприклад у У. crassa, гаплоїдні гетерокаріо- ни можна підтримувати необмежено довго. У інших грибів, наприклад у видів Aspergillus, цей процес далі розвивається в формі парасексу- ального циклу. Ядра гетерокаріонов іноді зливаються з утворенням диплоїдних ділянок міцелію, які можна селектировать при дії парів ^ -камфори, як це робиться для Aspergillus nidulans і Penicillium chrysogenum. Далі в ході мітотичних поділів диплоїдні ядра можуть зазнавати два незалежних процесу: гатоідізацію і митотичний крої- сінговеру - які ведуть до мітотичного або соматичному розщепленню.
Гаплоідізація відбувається спонтанно. Її ефективно індукує п-фторфеніл аланин.
Якщо при мітотичних діленнях диплоїдних ядер втрачається одна хромосома (2/7 - 1), то виникає анеуплоідних ядро стає нестабільним і послідовно втрачає все хромосоми одного набору, поки не встановиться стабільна гаплоидное число. При цьому хромосоми різних пар поводяться незалежно, а гени однієї хромосоми виявляють абсолютне зчеплення. У зв'язку з цим для локалізації невідомого гена в уже маркованої групі зчеплення необхідно встановити, з якими з маркерів постійно асоційований досліджуваний ген при гаплоідізаціі.
Локалізувати ген вже в межах групи зчеплення можна на основі мітотичного кросинговеру, спонтанного або індукованого реком- біногеннимі факторами. Кроссинговер на ділянці ген- центромера призводить до гомозиготизації всіх генів, розташованих дистальніше точки обміну в половині ядер (клітин) - нащадків рекомбінантного ядра (рис. 7.11). Таким чином, мітотичний Кросинговер можна використовувати для локалізації генів одного плеча хромосоми по відношенню до центромере.
При цьому відстань найбільш віддаленого від центромери маркера (гомозиготизації за яким відбувається найчастіше) приймають за 100%, а розташування інших маркерів визначають за формулою
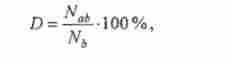
де D - відстань ген-центромера, Nb - загальне число мітотичних сегре- Гант, гомозиготних по b Nab - число сегрегантов, гомозиготних по а і b одночасно, якщо Комерсант - маркер, найбільш віддалений від центромери.
При вивченні рекомбінації з центромерой гени різних плечей однієї хромосоми не виявляється зчеплення, т. Е. Поводяться незалежно. Зчеплення між ними встановлюється при дослідженні гаплоідізаціі.
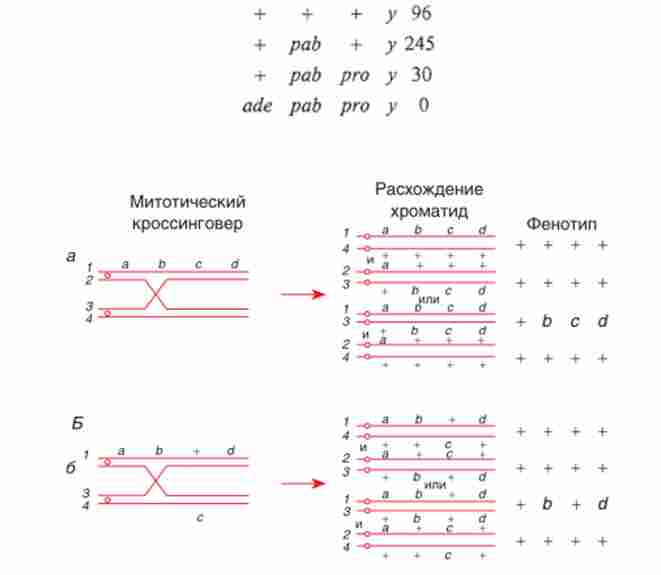
Розглянемо картування на основі мітотичного кросинговеру на прикладі даних, отриманих Г. Понтекорво і Г. Кейфер (1958) для Aspergillus nidulans.
Діплоїд

дав наступне співвідношення (серед 371 митотичних рекомбінантов, гомозиготних по у, який служив в даному випадку в якості маркера- селектора):
Мал. 7.11. Результат митотичного кросинговеру на ділянці а-b в тетрагетерозіготе по зчепленим генам, коли рецесивний аллель картіруемого гена з знаходиться: а - в маркірованому гомологів; б- в немаркованих гомологів;
«+» - домінантні аллели генів a, b, с, d
У попередніх дослідах по гаплоідізаціі було показано, що всі чотири гени знаходяться в одній групі зчеплення. Найбільшу відстань від центромери приймається за 100%; відстань центромера - /? ™ - 30: 371 х 100 % - 8%; центромера- /? А6 - 275: 371 х 100% = 74 %. Таким чином встановили порядок розташування генів і їх відстань від центромери (рис. 7.12). Рекомбінанти, гомозиготні по ade, не було отримано, можливо, через дуже тісного зчеплення ade з центромерой або через те, що ade знаходиться в іншому плечі хромосоми, а подвійний митотичний кроссинговер трапляється дуже рідко. Таким чином, парасексуаль- ний процес, незважаючи на те, що різні етапи його мають низьку частоту, дає надійні результати при генетичному аналізі.

Мал. 7.12. Розташування трьох генів по відношенню до центромере (ц) на основі мітотичного кросинговеру у Aspergillus nidulans
На картах груп зчеплення, побудованих на основі врахування мейотіче- ської і мітотичної рекомбінації, т. Е. При парасексуальними процесі, гени розташовуються в однаковій послідовності, як це показано для Aspergillus nidulans.