| Головна |
| «« | ЗМІСТ | »» |
|---|
За хімічною структурою всі гормони можуть бути розділені на кілька груп. Такий поділ особливо важливо, що передається гормонами інформація закодована в будові їх молекул.
Перша група - білково-пептидні гормони, що представляють собою амінокислотні ланцюжки різної довжини. До них відносяться ліберіни і статини гіпоталамуса, гормони гіпофіза, інсулін, натрійуретичний гормон і деякі інші.
Ці гормони сильно розрізняються за своїми розмірами: від олігопептидів, що складаються всього з трьох амінокислотних залишків (тіреотропін- рилізинг-фактор), до білків, що включають в себе до 2200 амінокислотних залишків (соматотропний гормон гіпофіза).
Білково-пептидні гормони синтезуються у вигляді великих білкових молекул - прегормони, які не володіють власною біологічною активністю. Як і інші білки, прегормони синтезуються на шорсткою ЕПР, де потім розщеплюються пептидазами до більш дрібних молекул - прогормонов - і в апараті Гольджі упаковуються в мембранні секреторні везикули. В процесі зберігання в цих везикулах прогормони розщеплюються, утворюючи деяку кількість активних молекул гормону. Гормони зберігаються в везикулах до тих пір, поки в організмі не виникає в них потреба (рис. 1.3).
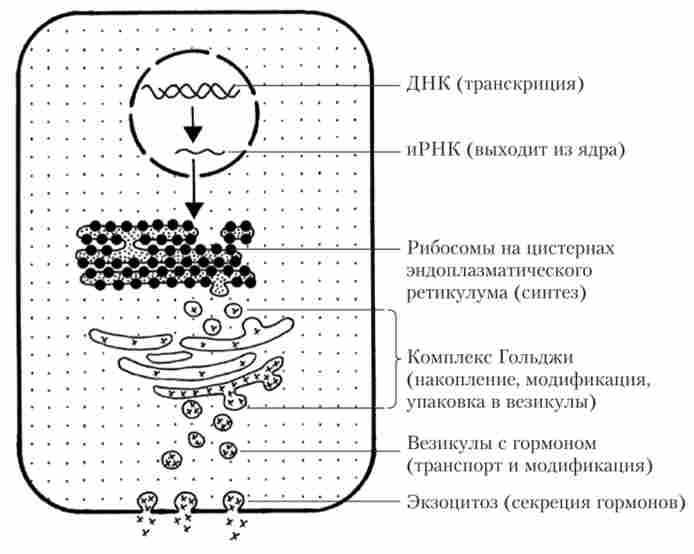
Мал. 1.3. Механізм утворення і секреції пептидних гормонів
Секреція гормонів здійснюється в тому випадку, коли везикули вбудовуються в клітинну мембрану шляхом екзоцитозу і що містяться в них гормони потрапляють у міжклітинний простір або безпосередньо в кровотік.
У багатьох випадках стимулом для екзоцитозу є збільшення концентрації іонів кальцію в цитоплазмі клітини, що викликається деполяризацией плазматичноїмембрани. В інших випадках стимуляція поверхневих рецепторів ендокринної клітини призводить до збільшення цАМФ і подальшої активації протеітсіназ, стимулюючих секрецію гормону.
Пептидні гормони водорозчинні (гідрофільних), що дозволяє їм легко проникати в кровоносну систему, що доставляє їх до тканям- мішенях, але не дозволяє вільно проходити через зовнішню клітинну мембрану, основу якої складають фосфоліпіди. Залози, секретіру- ющие білково-пептидні гормони, мають ентодермальні походження.
друга група - стероїди, що представляють собою похідні холестерину. До них відносяться гормони кори надниркових залоз (кортикостероїди, мінералокортикоїди) і статеві гормони (андрогени, естрогени і прогестини).
Стероїдні гормони жірорастворіми і складаються з трьох циклогексанового і одного циклопентанового кілець, об'єднаних в єдину структуру (рис. 1.4).
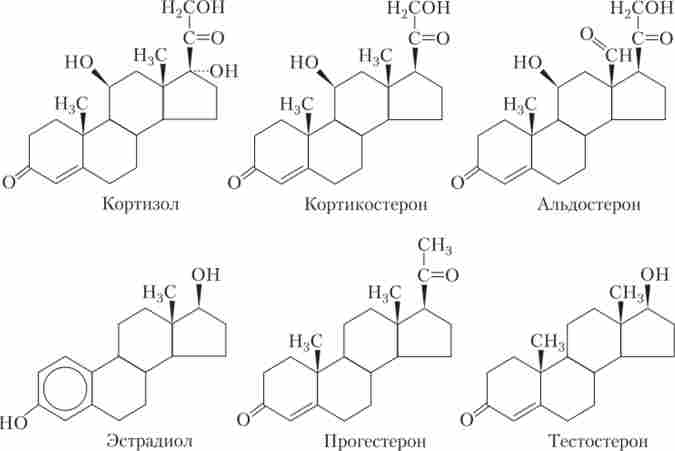
Мал. 1.4. Хімічні структури деяких стероїдів, що виробляються в кірковому шарі надниркових залоз і в статевих залозах
Зазвичай стероїди запасаються в секреторних клітинах в дуже малих кількостях, але зате в цих клітинах в вакуолях є запаси ефірів холе- стеролу, з яких в разі необхідності може бути синтезовано
достатню кількість гормонів. Холестерол надходить в клітини залоз з плазми крові. Так як стероїди ліпофільних, то вони легко дифундують через клітинні мембрани і потрапляють через міжклітинний простір в кровоносні капіляри. Досягнувши клітин-мішеней, стероїдні гормони легко проходять крізь їх зовнішню мембрану і зв'язуються зі своїми внутрішньоклітинними рецепторами. Стероідпродуцірующіе клітини мають мезодермальне походження.
третя група - гормони - похідні окремих амінокислот. Так, похідними амінокислоти тирозину є адреналін і НА - гормони мозкового шару надниркових залоз, а також гормони щитовидної залози - тироксин і трийодтиронін (рис. 1.5).
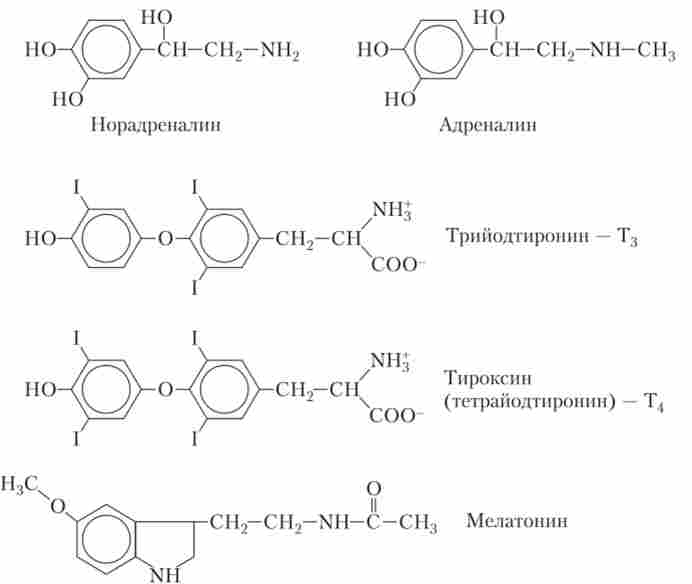
Мал. 1.5. Будова гормонів мозкового шару надниркових залоз, тиреоїдних гормонів щитовидної залози (Т3 і Т4) І гормону епіфіза мелатоніну
катехоламіни - адреналін і НА - синтезуються в клітинах мозкової речовини надниркових залоз, причому адреналіну синтезується в чотири рази більше, ніж НА. Ці гормони депонуються в мембранних бульбашках до моменту стимуляції клітини і секреції шляхом екзоцитозу. В кровотоці адреналін і НА можуть бути як у зв'язаному, так і у вільному стані.
тиреоїдні гормони синтезуються і зберігаються в щитовидній залозі, включаючись в макромолекули білка тиреоглобуліну, який розміщується у великих фолікулах щитовидної залози. Секреція гормону здійснюється за умови відщеплення амінів від тиреоглобуліну, вільні гормони виділяються в кровотік. Після потрапляння в кров велика частина молекул тиреоїдних гормонів зв'язується з тирозин-глобуліном (білком-переносником) і частково з деякими іншими білками плазми. При необхідності гормони отщепляются від цих білків і взаємодіють з внутрішньоклітинними рецепторами.
мелатонін - ще один гормон, що відноситься до цієї хімічної групи, - секретується епіфізом. Мелатонін синтезується з амінокислоти триптофану.
четверта група - простагландини, тромбоксани, лейкотрієни - похідні арахідонової кислоти, що відноситься до класу ненасичених жирних кислот. Вони були виявлені в багатьох тканинах людини та інших ссавців. Раніше ці речовини відносили до гістогормонам, що володіє місцевою дією, проте до теперішнього часу доведено, що ці речовини здатні вступати до кровотік і надавати системний вплив на функції організму. Простагландини синтезуються шляхом циклізації ділянки в центрі вуглеводневого ланцюга (С20) ейкозанових поліненасичених кислот, частіше арахідонової (рис. 1.6).
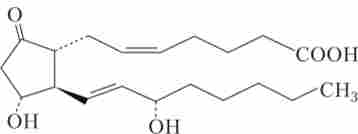
Мал. 1.6. Будова молекули простагландину Е2 (PgE2)
Родинна група регуляторів - тромбоксани, виділені спочатку з тромбоцитів; містять Циклопентанова кільце, в яке включений атом кисню.
Класифікація гормонів за хімічною структурою представлена в табл. 1.3.
Таблиця 1.3
Класифікація гормонів за хімічною структурою
|
клас |
сімейство |
ряд |
представники |
|
стероїдні |
З21-стсроіди (Прегнановие) |
кортикостероїди: глюкокортикоїди |
Кортікостсрон, гідрокортизон |
|
кортикостероїди: мінералокортикоїди |
альдостерон |
||
|
Прогесгіни |
прогестерон |
||
|
З19-стероїди (Андростановие) |
андрогени |
тестостерон; 5а-дигідротестостерон |
|
|
З18-стероїди (Естрановие) |
естрогени |
Естрадіол, естрон, естріол |
|
|
З27-стероїди (Холестановие) |
вітамін D:) |
l, 25 (OH)2D3 |
|
клас |
сімейство |
ряд |
представники |
|
Похідні іена- насичення жирних кислот |
ретиноїди (С20: 5) |
ретиноевую кислоти |
ретиноєва кислота |
|
ейкозаноїди
|
тромбоксани |
ТХА2 |
|
|
простагландини |
PGF2a |
||
|
Простаці кліпи |
PGI{ |
||
|
лейкотрієни |
lta4 |
||
|
похідні амінокислот |
похідні тирозину |
катехоламіни |
Адреналін, дофамін |
|
тиреоїдні гормони |
Тироксин, трійодтіро- нин |
||
|
похідні триптофану |
мелатонін |
мелатонін |
|
|
Серотонін (гістогормон) |
серотонін |
||
|
білково пептидні |
Пептиди нейро- гіпофіза |
ряд вазопресину |
Аргінін-вазопресин |
|
ряд окситоцину |
окситоцин |
||
|
Рилізинг-фак- тори гіпоталамуса |
кортіколіберін, соматостатін |
||
|
ангіотензин |
ангіотензин II |
||
|
олігопептиди АКТГ-типу |
АКТГ, а-МСГ, р-ліпотропін |
||
|
Олігопептиди шлунково-кишкового тракту |
Ряд глюкагону. Ряд гастрину. Ряд гастрин-рилізинг пептиду. Ряд соматостатина. Ряд панкреатичного поліпептиду |
Глюкагон, секретин, вип. Гастрін, холецістокі- нин. ГРП, субстанція Р. Нейротензин. нейропептид Y |
|
|
сімейство інсуліну |
Інсулін і інсуліноподібний ростові фактори |
інсулін, IGF-l, IGF-U |
|
|
релаксин |
релаксин |
||
|
гормони тимуса |
Тимозин, тимопоетин |
||
|
Атріопентіди |
натрийуретичні чинники ANF, BNF |
||
|
Олігопептід- ні регулятори обміну Са2^ |
ряд кальцитонина |
Кальцитонін, кальці- тонин ген-роственний пептид, адреномедуллін |
|
|
ряд паратгормона |
Паратгормон, білок, споріднений паратгормону |
|
клас |
сімейство |
ряд |
представники |
|
Мономірні білки типу СТГ |
Соматотропін, про- лактин, плацентарний лактоген |
||
|
Мономірні білки типу FGF (Гістогормони) |
FGR3, EGF |
||
|
Цитокіни (гисто- |
інтерлейкіни |
IL-, IL-3, еритропоетин |
|
|
гормони) |
інтерферони |
INFa, INFy |
|
|
лептин |
лептин |
||
|
Димерні глікопротеїди типу Л Г |
Л Г, хоріонічний гонадотропін, тиреотропин |
||
|
Димерні глікопротеїди типу TGF3 |
Фактор регресії мюл- леров каналів, акти- провини, ингибин, TGF |
||
|
Димерні білки типу фактора росту тромбоцитів (гістогормони) |
PDGF, VEGF |
Незважаючи на величезну різницю в хімічному будову, гормони різних хімічних груп мають багато спільного в організації функціональної структури.
Так, в ідеальному випадку в молекулі гормону можна виділити адресний ділянку, або гаптон, відповідальний за взаємодію всієї молекули з місцем специфічного гормонального впливу. Гаптон сам по собі жодним біологічним ефектом не володіє. Інша ділянка гормональної молекули називається Актон. Саме Актон викликає фізіологічні ефекти гормону в клітці-мішені, але сам по собі без гаптона Актон не може приєднуватися до регульованої клітці. І нарешті, до складу гормональної молекули може входити цілий ряд ділянок, які захищають цю молекулу від дії руйнівних ферментів, що підсилюють або послаблюють гормональний ефект. Ці ділянки молекули отримали назву акцесорних. Актон, гаптон і акцесорних ділянки можуть бути розташовані в різних частинах гормональної молекули, а можуть і перекривати один одного (рис. 1.7).
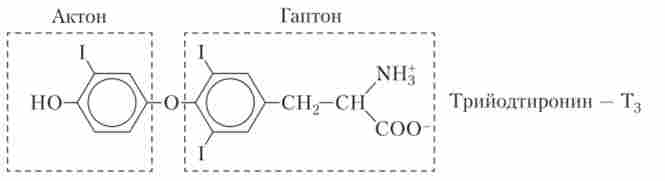
Мал. 1.7. Розташування Актон і гаптонного ділянок в молекулі
трийодтиронина (Т3)
Потрапивши в кров, багато гормони специфічно зв'язуються з транспортними білками. Пов'язаний зі своїм транспортним білком гормон вимикається з сфер біологічної дії і метаболізму, за рахунок чого знижується швидкість його розпаду, і збільшується період його напіввиведення. Це дозволяє створювати певний депо гормону, пом'якшуючи різкі зміни його концентрації в крові. В даний час відкрито багато транспортних білків, що продукуються, головним чином, печінкою, які пов'язують гормони в крові. Детальніше транспорт гормонів описаний в параграфі 1.7.