| Головна |
| «« | ЗМІСТ | »» |
|---|
Центральною подією гаметогенеза є особлива форма клітинного ділення - мейоз. На відміну від широко поширеного мітозу, що зберігає в клітинах постійне диплоидное число хромосом, мейоз призводить до утворення з диплоїдних клітин гаплоїдних гамет. При подальшому заплідненні гамети формують організм нового покоління з диплоїдним кариотипом (пс + пс = = 2п2с). У цьому полягає найважливіше біологічне значення мей- оза, який виник і закріпився в процесі еволюції у всіх видів, що розмножуються статевим шляхом (див. Розд. 3.6.2.2).
Мейоз складається з двох швидко наступних один за іншим поділів, що відбуваються в періоді дозрівання. Подвоєння ДНК для цих поділів здійснюється одноразово в періоді зростання. Друге поділ мейозу слід за першим практично відразу так, що спадковий матеріал не синтезується в проміжку між ними (рис. 5.5).
Перше мейотичне розподіл називають редукційним, так як воно призводить до утворення з диплоїдних клітин (2п2с) гаплоїдних клітин п2с. Такий результат забезпечується завдяки особливостям профази першого поділу мейозу. У профазі I мейозу, так само як в звичайному мітозі, спостерігається компактна упаковка генетичного матеріалу (спирализация хромосом). Одночасно відбувається подія, якого немає у мітозі: гомологічні хромосоми кон'югують один з одним, т. Е. Тісно зближуються відповідними ділянками.
В результаті кон'югації утворюються хромосомні пари, або біваленти, числом п. Так як кожна хромосома, що вступає в мейоз, складається з двох хроматид, то бивалент містить чотири хро- матіди. Формула генетичного матеріалу в профазі I залишається 2п4с. До кінця профази хромосоми в бівалентах, сильно спіралізуясь, коротшають. Так само як в мітозі, в профазі I мейозу починається формування веретена поділу, за допомогою якого хромосомний матеріал буде розподілятися між дочірніми клітинами.
Процеси, що відбуваються в профазі I мейозу і визначають його результати, обумовлюють більш тривалий перебіг цієї фази розподілу в порівнянні з митозом і дають можливість виділити кілька стадій в її межах.
Лептотена - найбільш рання стадія профази I мейозу, в якій починається спирализация хромосом і вони стають видимими
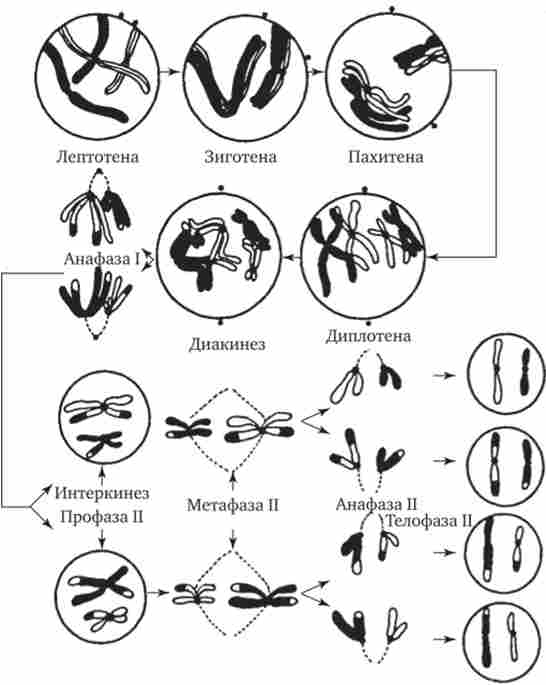
Мал. 5.5. Стадії мейозу.
Батьківські хромосоми позначені чорним кольором, материнські - нефарбовані. На малюнку не зображена метафаза I, в якій біваленти розташовуються в площині екватора веретена поділу, і телофаза I, швидко переходить в профазу II

в мікроскоп як довгі і тонкі нитки. Зиготена характеризується початком кон'югації гомологічних хромосом, які об'єднуються сінаптонемальним комплексом в бивалент (рис. 5.6). Пахитена - стадія, в якій на тлі триваючої спирализации хромосом і їх укорочення між гомологічними хромосомами здійснюється кроссинговер - перехрещення з обміном відповідними ділянками. Диплотена характеризується виникненням сил відштовхування між гомологічними хромосомами, які починають віддалятися один від одного в першу чергу в області центромер, але залишаються пов'язаними в областях минулого кросинговеру - хіазмі (Рис. 5.7).

Мал. 5.8. Стадія діакінеза в мейозі у людини.
Стрілками вказані хіазми

Мал. 5.7. Стадії діплотени в мейозі коника
діакінеза - завершальна стадія профази I мейозу, в якій гомологічні хромосоми утримуються разом лише в окремих точках хиазм. Біваленти набувають химерних форм кілець, хрестів, вісімок і т. Д. (Рис. 5.8).
Таким чином, незважаючи на виникаючі між гомологічними хромосомами сили відштовхування, в профазі I не відбувається остаточного руйнування бівалентов. Особливістю мейозу в овогенезі є наявність спеціальної стадії - діктіотени, відсутньої в сперматогенезе. На цій стадії, що досягається у людини ще в ембріогенезі, хромосоми, прийнявши особливу морфологічну форму «лампових щіток», припиняють будь-які подальші структурні зміни на багато років. Після досягнення жіночим організмом репродуктивного віку під впливом лютеїнізуючого гормону гіпофіза, як правило, один овоціт щомісяця вступає в мейоз.
В метафазі I мейозу завершується формування веретена поділу. Його нитки прикріплюються до Центромера хромосом,
об'єднаних в біваленти, таким чином, що від кожної центромери йде лише одна нитка до одного з полюсів веретена. В результаті нитки, пов'язані з центромерами гомологічниххромосом, прямуючи до різних полюсів, встановлюють біваленти в площині екватора веретена поділу.
В анафазе I мейозу послаблюються зв'язки між гомологічними хромосомами в бівалентах і вони відходять один від одного, прямуючи до різних полюсів веретена поділу. При цьому до кожного полюсу відходить гаплоїдний набір хромосом, що складаються з двох хроматид (див. Рис. 5.5).
В телофазе I мейозу у полюсів веретена збирається одинарний, гаплоїдний набір хромосом, кожна з них містить подвійну кількість ДНК. Формула генетичного матеріалу утворюються дочірніх клітин відповідає п2с.
друге мейотічне (екваціонное) поділ призводить до утворення клітин, в яких вміст генетичного матеріалу в хромосомах буде відповідати їх однонитчатим структурі пс (Див. Рис. 5.5). Цей поділ протікає, як мітоз, тільки клітини, що вступають в нього, несуть гаплоїдний набір хромосом. У процесі такого поділу материнські двунитчатую хромосоми, розщеплюючи, утворюють дочірні однонитчатим.
Одна з головних задач мейозу - створення клітин з гаплоїдний набором однонитчатим хромосом - досягається завдяки одноразовій редуплікаціі ДНК для двох послідовних поділів мейозу, а також завдяки утворенню на початку першого мейотичного поділу пар гомологічних хромосом і подальшого їх розбіжності в дочірні клітини.
Процеси, які відбуваються в редукційний поділ, забезпечують також не менш важливе наслідок - генетичну різноманітність гамет, утворених організмом. До таких процесів відносять кроссін- Говер, розбіжність гомологічних хромосом в різні гамети і незалежна поведінка бівалентов в першому мейотичному поділі (Див. Розд. 3.6.2.3).
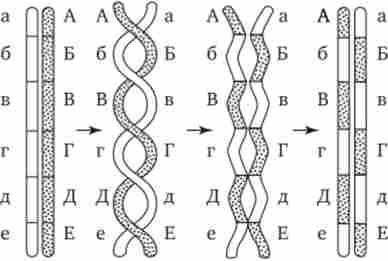
кроссинговер забезпечує перекомбінація батьківських і материнських алелів в групах зчеплення (див. рис. 3.72). З огляду на те що перехрещення хромосом може відбуватися в різних ділянках, кроссинговер в кожному окремому випадку призводить до обміну різними за кількістю генетичним матеріалом. Необхідно відзначити також можливість виникнення кількох перекрестов між двома хроматидами (рис. 5.9) і участі в обміні більш ніж двох хроматид бивалента (рис. 5.10). Зазначені особливості кросинговеру роблять цей процес ефективним механізмом перекомбінації алелей.
Розбіжність гомологічниххромосом в різні гамети в разі гетерозиготності приводить до утворення гамет, що розрізняються по аллелям окремих генів (див. рис. 3.74).
Випадкове розташування бівалентов в площині екватора веретена поділу і подальше їх розбіжність в анафазе I мейозу
Мал. 5.9. Багаторазовий кроссинговер між гомологічними хромосомами. А - Е, а - е - локуси хромосом
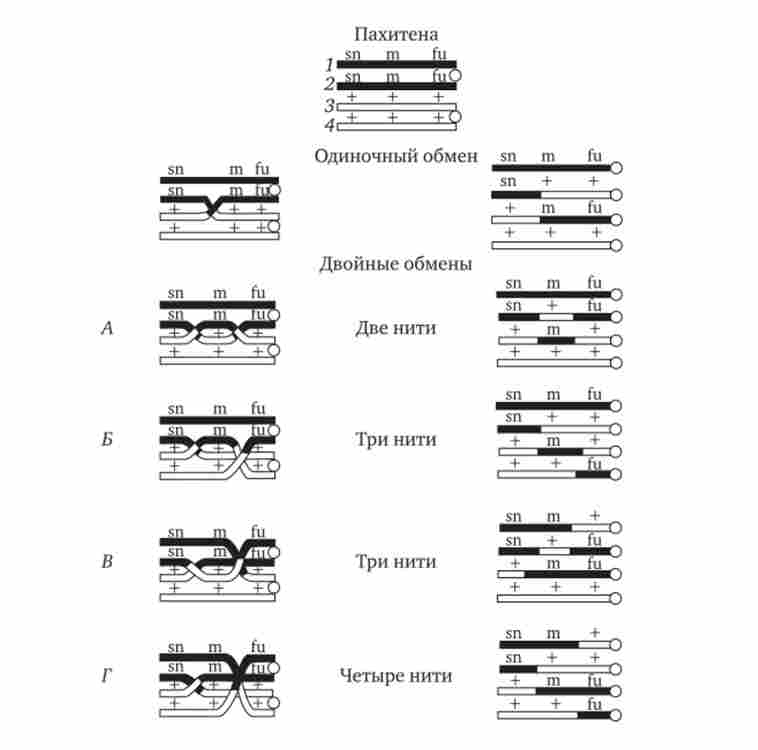
Мал. 5.10. Множинний обмін ділянками на стадії чотирьох хроматид в пахі- тене мейозу.
У кроссинговер можуть вступати всі чотири хроматиди бивалента, латинськими буквами позначені мутантні аллели; знаком «+» - нормальні аллели забезпечують рекомбінацію батьківських груп зчеплення в гаплоидном наборі гамет (див. рис. 3.75).
Останні стадії овогенеза відтворюються і поза організмом жінки, в штучному живильному середовищі. Це дозволило здійснити зачаття людини «в пробірці». Перед овуляцією хірургічним шляхом яйце витягується з яєчника і переноситься в середу зі сперматозоїдами. Що виникає в результаті запліднення зигота, будучи поміщена в сприятливе середовище, здійснює дроблення. На стадії 8-16 бластомерів зародок переноситься в матку жінки-реципієнта, яка здійснює доношування і пологи. Кількість проданих результатів такого перенесення останнім часом зростає.
Гаметогенез відрізняється високою продуктивністю. За час статевого життя чоловік продукує не менше 500 млрд сперматозоїдів. На п'ятому місяці ембріогенезу в зародку жіночої статевої залози налічується 6-7 млн клітин - попередниць яйцеклітин. До початку репродуктивного періоду в яєчниках виявляється приблизно 100 000 овоцитов. Від моменту статевого дозрівання до припинення гаметогенеза в яєчниках дозріває 400-500 овоцитов.