РЕАКЦІЇ ОКИСЛЕННЯ, НЕ ПОВ'ЯЗАНІ З ДІЄЮ МОНООКСИГЕНАЗ
Деякі ферментні системи здатні окисляти ксенобіотики. До них відносяться:
флавінсодержащіе монооксідази, локалізовані в мікросомах, каталізують окислення вторинних і четвертинних амінів, гидразинов, сірку і фосфоровмісних сполук;
алкоголь-і альдегіддегідрогеназние системи, локалізовані в мікросомах і цитозолі. У присутності НАДФН і кисню окислюють етанол до альдегіду і оцтової кислоти відповідно;
пероксидазного-каталазна системи окислюють органічні субстрати, а також беруть участь в окисленні етанолу;
ксантиноксидаза каталізує окислення ксантину і ксантінсодержа- ших речовин до сечової кислоти;
амінооксідаза каталізує окислення амінів до відповідних альдегідів;
окислительное дегалогенізірованіе, пов'язане із заміщенням хлору на кисень, наприклад при перетворенні ДДТ в ДДА.
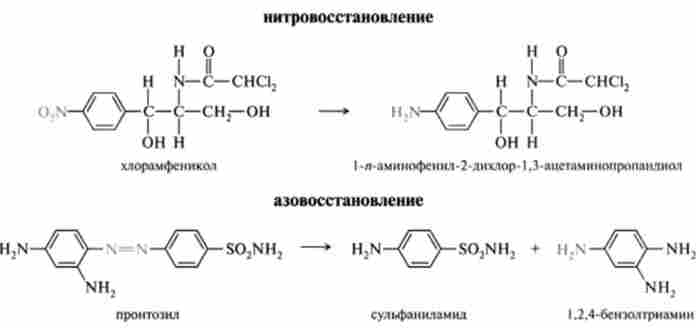
Реакції відновлення в ЕПР протікають за участю НАДФН-залежного флавопротеїнів і цитохрому Р-450. Найбільш часто зустрічається відновлення нітро- і азосоединений:
НАДН-залежні реакції вносять значно менший внесок в биотрансформацию ксенобіотиків. У мікросомах локалізовані принаймні дві ферментні системи, здатні відновлювати ксенобіотики. Обидві системи - флавопротсіни з ФАД як про- стетичне групи. Класичним прикладом мікросомального відновлення є перетворення нітробензолу в анілін.
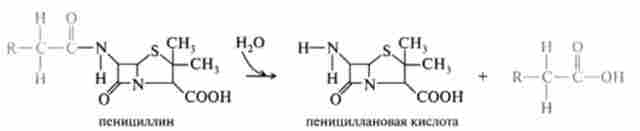
Найбільш поширеними реакціями гідролізу ксенобіотиків є естеразной реакції. Вони грають дуже важливу роль у розвитку токсичного ефекту при гідролізі фосфорорганічних речовин. Можна також відзначити гідроліз під дією неспецифічних естераз складних ефірів, амідів, гідроксамових кислот, гідразидів та ін. Наприклад:
Реакція біоактивації ксенобіотиків часто призводить до змін в молекулі ксенобиотика, які збільшують її полярність, розчинність і сприяють швидшому виведенню з організму. Так як токсичність в чималій мірі обумовлена накопиченням речовини в клітинах, можна вважати, що виведення вихідного ксенобиотика або його метаболітів з організму призводить до зменшення або повного зникнення токсичного ефекту. Однак існує багато винятків, наприклад, в ряді реакцій утворюються продукти з більшою токсичністю в порівнянні з вихідною речовиною, особливо в реакціях першої фази системи біотрансформації.
Це не дивно, так як впровадження в молекулу полярної групи може значно збільшити її реакционноспособна по відношенню до інших сполук.
Реакції біотрансформації, в яких утворюються продукти, що мають велику токсичність у порівнянні з вихідним ксенобіотиком, називаються реакціями біоактивації. Існує багато прикладів подібних реакцій, причому найбільш відома реакція перетворення інсектициду паратіона в па- раоксон. Маратом належить до органотіофосфатам, нейротоксичність яких заснована на їх взаємодії з ферментом ацетилхолінестеразою. Спорідненість цього ензиму до параоксон у багато разів вище, ніж до вихідного з'єднанню - паратіон. Іншими словами, реакція окислення, необхідна для перетворення речовини в більш розчинний з'єднання, призводить до утворення продукту біоактивації.
В результаті біоактивації можуть утворюватися токсичні інтермедіати, що індукують патологічні процеси в організмі (табл. 32.1).
Таблиця 32.1. Токсична дія метаксенобіотіков на організм
речовина
Формула
метаболіт
ефект
бромбензил
-Про
чу
некроз печінки
анілін
h,nhQ
НО-NH-
Мстгсмоглобінсмія
Д і мстив і ітроза м і і
H} C^> N-N = 0 Н3З ^
Н, (Г
канцерогенез
вінілхлорид
нз = зн
H ' ^ Cl
нон
х-х
н про
рак печінки
Регуляція мітохондріального окислення - біохімія Факторами, що лімітують швидкість дихання, є доступність кисню, АДФ, субстратів, можливості і стан самої дихального ланцюга при насичують концентраціях всіх субстратів і компонентів. При наявності всіх компонентів дихального ланцюга і субстратів, за винятком АДФ, поглинання 0 2 (Дихання) не
Регуляція кровообігу, загальні положення - фізіологія людини і тварин В результаті вивчення даного розділу студенти повинні: знати особливості забезпечення організмом кровио всіх органів і тканин в точній відповідності з мінливими потребами; характер взаємодії місцевих, нервових і гуморальних регуляторних механізмів; координацію роботи серцево-судинної системи
Регуляція експресії генів у прокаріотів - біологія. Частина 1 Вивчення регуляції генної активності у прокаріотів призвело французьких мікробіологів Ф. Жакоба і Ж. Моно до створення (1961) оперон моделі регуляції транскрипції. оперон - це тісно пов'язана послідовність структурних генів, що визначають синтез групи білків, які беруть участь в одного ланцюга
Регуляція біосинтезу гема - біохімія частина 2. Основний швидкістьлімітувальною реакцією синтезу гема є конденсація гліцину і сукцинил-КоА, що каталізується амінолевулінатсінтазой (АЛ-синтаза). Встановлено, що в рстікулоцітах регуляція здійснюється на рівні синтезу АЛ-синтази на стадії ініціації трансляції. При цьому Мал. 25.3. Схема регуляції
Регуляторні пептиди - фізіологія людини і тварин Серед сигнальних молекул, що беруть участь в хімічній передачі інформації, важливе місце належить РП. Ці речовини є полімерні ланцюжки, що містять від 2 до 70 амінокислотних залишків. Більші молекули такого типу, що володіють сигнальної функцією, класифікують як регуляторні білки. РП синтезуються
Рефлекторні дуги спинного мозку - анатомія центральної нервової системи Для того щоб зрозуміти, чому утворено і як функціонує сіра і біла речовина СМ, треба знати, як влаштовані дуги спинномозкових рефлексів. При цьому потрібно враховувати, що за участю СМ здійснюються найрізноманітніші реакції, в тому числі довільні рухи, керовані ГМ. Однак існують і відносно