| Головна |
| «« | ЗМІСТ | »» |
|---|
Для того щоб зрозуміти, чому утворено і як функціонує сіра і біла речовина СМ, треба знати, як влаштовані дуги спинномозкових рефлексів. При цьому потрібно враховувати, що за участю СМ здійснюються найрізноманітніші реакції, в тому числі довільні рухи, керовані ГМ. Однак існують і відносно прості безумовні (т. Е. Що йдуть по вродженої заданою програмою) рефлекси СМ, які він здатний реалізовувати самостійно. За допомогою СМ здійснюються безумовні рефлекси скелетної мускулатури тіла (соматичні рефлекси) і безумовні рефлекси внутрішніх органів (Вегетативні рефлекси).
Розглянемо рефлекторну дугу СМ на прикладі соматичного колінного рефлексу (рис. 5.4, а). Це найпростіший безумовний рефлекс, в дугу якого, як уже зазначалося вище (див. Параграф 3.3), входять тільки два нейрона - сенсорний і руховий. Такі рефлекси називають моно- сінантічними, так як в дузі рефлексу присутній тільки один розташований в ЦНС (центральний) синапс між нейронами; Наступного синапс - вже нервово-м'язовий.
Спинномозкові ганглії в дорсальних корінцях СМ утворені скупченням псевдоуніполярних сенсорних нейронів. Такі нейрони мають дуже довгий периферичний відросток, який функціонально є дендритом. Периферичні відростки псевдоуніполярних нейронів проводять інформацію від самих різних рецепторів - шкірних (больових, тактильних, температурних), пропріорецепторов (м'язово-суглобових), вісцерорецеіторов (рецепторів внутрішніх органів). При реалізації колінного рефлексу кінцеві розгалуження такого дендрита збуджуються при розтягуванні чотириголового м'яза стегна в результаті удару по її сухожилля. Нервовий імпульс (ПД) по дендрити проводиться до тіла чутливої клітини і далі по відносно короткому аксону в СМ. У вентральном розі сірої речовини цей аксон утворює синапс на руховому виконавчому нейроне СМ, викликаючи його збудження. У свою чергу ПД по аксону мотонейрона доходить до розтягнутої м'язи і через нервово-м'язовий синапс запускає її скорочення. В результаті м'яз коротшає, і колінний суглоб розгинається. Подібні рефлекси можна отримати при розтягуванні будь-який інший м'язи нашого тіла.
Дуги більш складних рефлексів (наприклад, згинання руки у відповідь на больовий стимул) мають кілька интернейронов в задніх рогах і в проміжному речовині СМ. Такі рефлекси називають полісинаптичні
(Рис. 5.4, а). Наявність интернейронов в рефлекторних дугах істотно збільшує час реакції таких рефлексів, так як передача імпульсу через синапс здійснюється з деякою затримкою. Але ГМ здійснює регуляцію рефлексів головним чином саме через вставні нейрони, завдяки чому стає можливим довільний контроль полісінап- тичних рефлексів.
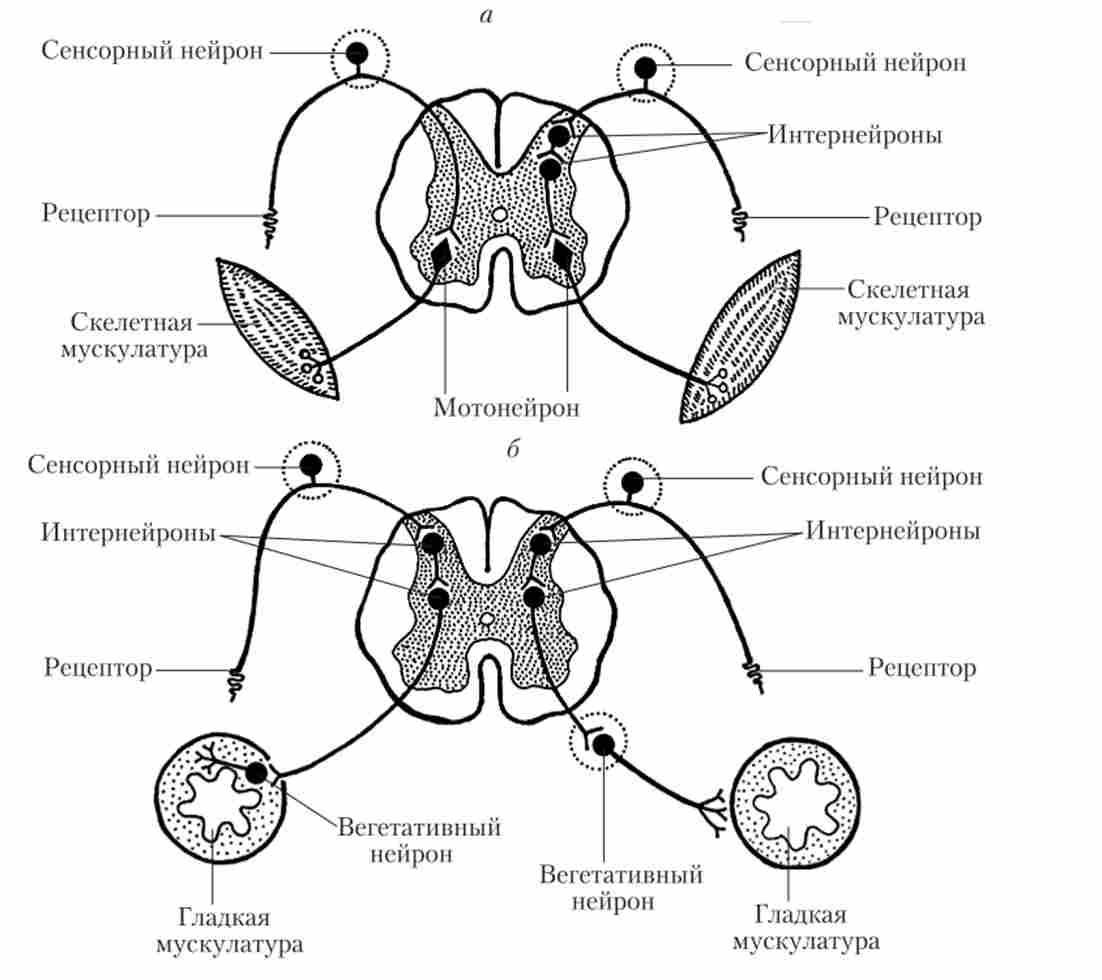
Мал. 5.4. Рефлекторні дуги спинного мозку:
а - дуги соматичних рефлексів (зліва - моносинаптічеськие, праворуч - полісинаптичні); б - дуги вегетативних рефлексів (зліва - парасимпатичного,
праворуч - симпатичного)
Обидві розглянуті вище рефлекторні дуги є дугами соматичних рефлексів (рефлексів скелетної мускулатури). Дуги вегетативних рефлексів помітно відрізняються від соматичних (див. Рис. 5.4, б). Основна відмінність полягає в місцезнаходження виконавчого нейрона в рефлекторну дугу. Цей нейрон розташований не в ЦНС (як в соматичної НС), а в периферичному вегетативному ганглії. чутливі
нейрони автономної НС знаходяться там же, де і чутливі нейрони соматичної НС - в спінальних гангліях на задніх корінцях спинного мозку. Потім може слідувати ланцюжок вставних центральних нейронів. Останній (а іноді - єдиний) центральний нейрон вегетативної дуги в СМ розташований або в бічних рогах сірої речовини (симпатичні нейрони), або в проміжному речовині крижового відділу (парасимпатичні нейрони). Аксон цього центрального вегетативного (пре- гангліонарного) нейрона прямує до нейронам вегетативного ганглія, які іннервують виконавчий орган. Більшість симпатичних гангліїв знаходяться поруч з СМ, парасимпатичні ганглії лежать або поруч з иннервируемой органом, або в його стінках.
Після вивчення пристрою дуг спинномозкових рефлексів стає зрозуміло, чому утворені як задні і передні корінці, так і спинномозкові нерви. Аксони псевдоуніполярних сенсорних нейронів спінальних гангліїв входять в СМ, утворюючи спинні (чутливі) корінці. Частина цих аксонів закінчується (утворює синапси) на вставних нейронах або мотопейронах, інша частина в складі білої речовини піднімається в ГМ. Дендрити (периферичні відростки) сенсорних нейронів йдуть у складі відповідного спинномозкового нерва на периферію, утворюючи там чутливі нервові закінчення. вентральні (рухові) корінці утворені аксонами мотонейронів і прегангліонаріих вегетативних нейронів.
спинномозкові нерви утворюються в результаті об'єднання передніх і задніх корінців, тому вони є змішаними, гак як утворені волокнами двох типів - аферентні (доцентровими) сенсорними і еферентних (відцентровими) виконавчими, т. е. що йдуть до різного типу м'язів і залоз.
Відразу після виходу з хребетного каналу спинномозкові нерви поділяються на передню і задню гілки, в кожну з яких входять як чутливі, так і рухові волокна. Надалі ці гілки поділяються на все більш дрібні периферичні нерви.
Ділянка шкіри, іннервіруємий однією парою спинномозкових нервів, називають дерматомом, а сукупність м'язів, які іннервує ця пара, називається міотомія. Але якщо подивитися на обриси дерматомов і міотомів на тілі людини, стає очевидно, що їх розташування не завжди відповідає місцезнаходженню сегмента (рис. 5.5). Пов'язано це з тим, що під час ембріонального розвитку спочатку кожному сегменту СМ чітко відповідають свій дерматом і миотом.
Однак надалі, по-перше, окремі м'язи можуть утворюватися з декількох сусідніх миотомов і, відповідно, бути пов'язаними з декількома сусідніми корінцями; і, по-друге, завдяки існуванню сплетінь (див. параграф 3.1) волокна від одного корінця в процесі розгалуження потрапляють до складу декількох нервів, тому на кордоні сусідніх сегментів є ділянки перекриття зон іннервації. В результаті, якщо порівнювати клінічну картину при ураженні периферичного нерва і при ураженні корінців, від яких цей нерв отримує волокна, вона буде різна.
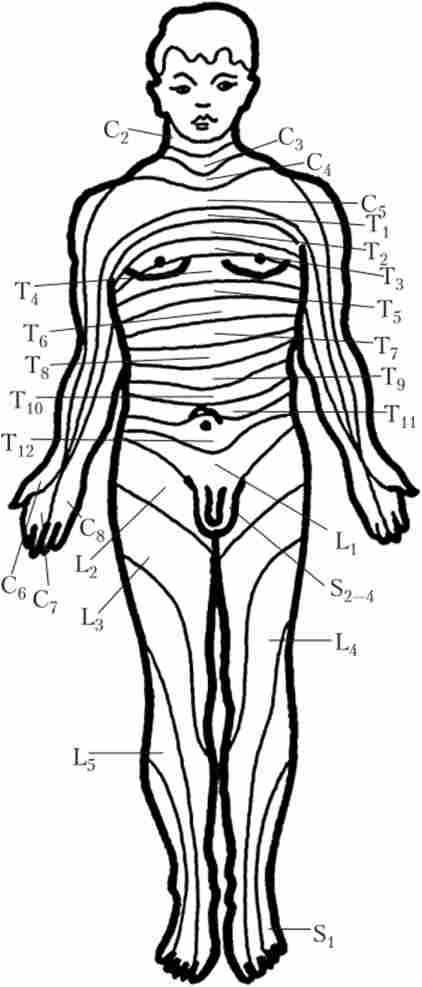
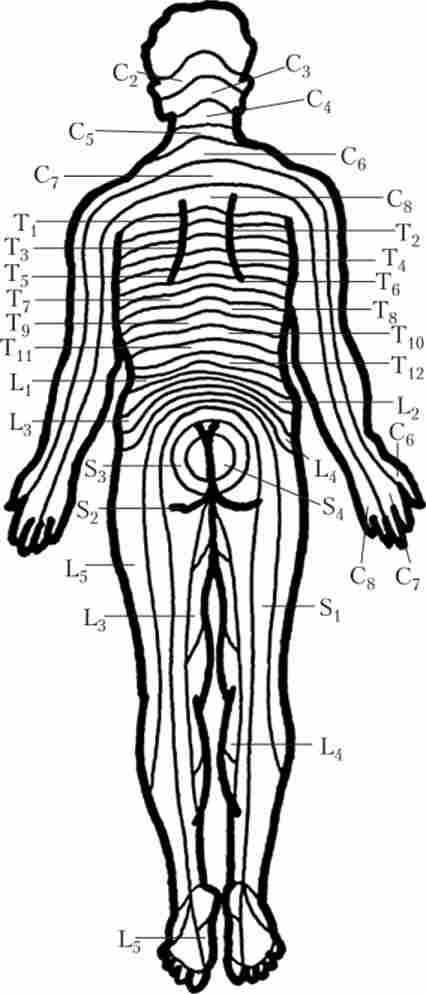
Мал. 5.5. дерматоми людини
Іннервація внутрішніх органів носить ще більш складний характер, так як більшість цих органів иннервируется не одним сегментом СМ, а декількома, до того ж одними і тими ж для ряду органів, наприклад, тонкий кишечник - від сегментів Т9-Т12, нирки - від Т10-L2. Тому не завжди просто при виникненні больових відчуттів виявити уражений орган, так як біль носить не строго обмежений, а розлитої характер. Можливо навіть поява відображеної болю, т. Е. Болю, яка не співпадає з місцем свого виникнення з локалізацією патологічного процесу.
У зв'язку з цим велике практичне значення має знання зон Захар'їна - Геда. Це ділянки шкіри, в яких при захворюваннях певних внутрішніх органів можуть виникати відбиті болю (рис. 5.6).
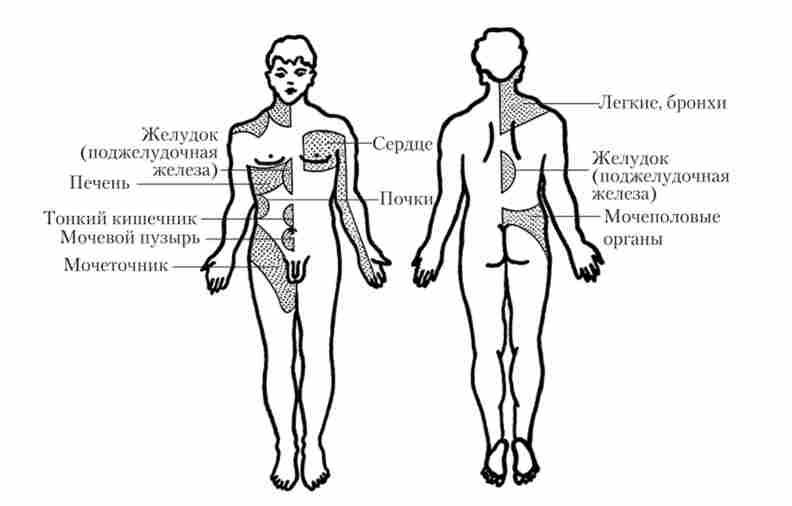
Мал. 5 6. Зони Захар'їна - Геда (відображення болю при захворюваннях внутрішніх органів)
Можливо і поява на них гіперестезії, т. З. підвищеної чутливості до подразників. Для багатьох внутрішніх органів зони Захар'їна - Геда встановлені досить точно. Так, наприклад, добре відома гіперестезія в правій клубової області при апендициті. Є припущення, що наявність цих зон пояснює лікувальну дію різних методів рефлексотерапії (голковколювання, припікання і т. П.).