
| Головна |
| «« | ЗМІСТ | »» |
|---|
Сітківка у ссавців тварин складається з декількох шарів клітин різного типу. Перш ніж досягти фоторецепторних клітин, світлові промені повинні пройти через всі шари цих клітин (див. Рис. 15.16, Б). Фоторецепторні клітини - це сильно видозмінені волоскові клітини. Сома клітини - внутрішній сегмент містить ядро і велику кількість дрібних мітохондрій (рис. 15.17, А); в базальної частини є афферентное синаптічеськоє закінчення, а від верхнього кінця відходить рудиментарная ресніч-
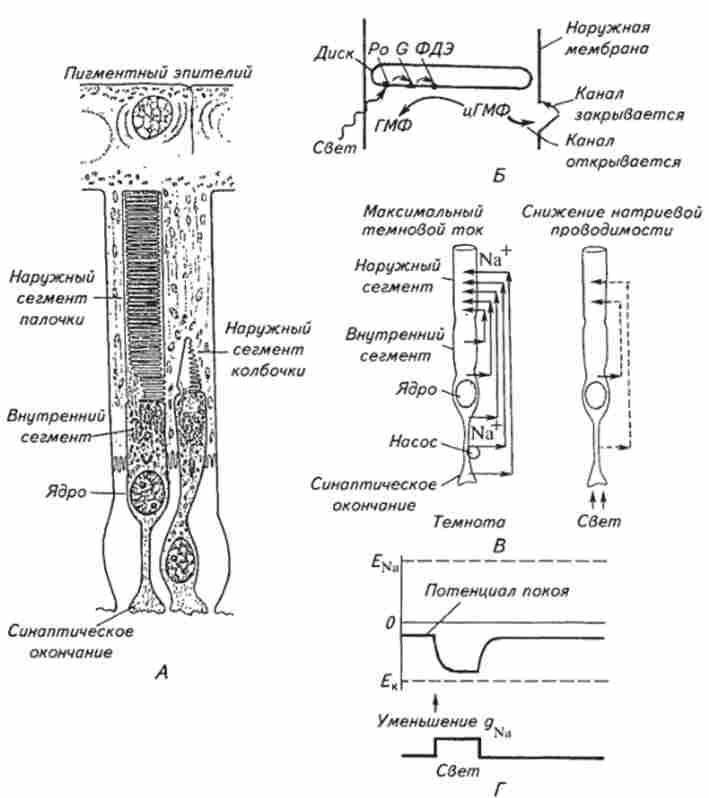
Мал. 15.17. Морфофункціональна організація фоторецепторів хребетних
тварин:
А. Паличка і колбочка сітківки. Б. Схема передачі фотозбудження до іонним каналам зовнішньої мембрани рецептора: Р0 - фотопігмент родопсин; G- білок; ФДЕ - фосфодіхте- рази; ГМФ - гуанозінмонофос<1> ат; іГМФ - циклічний гуанозинмонофосфат. В Зміна іонного струму через мембрану фоторецептора при висвітленні. Г. Зміна мембранного потенціалу (потенціалу спокою) фоторецептора при висвітленні?Na - рівноважний потенціал для іонів натрію, Едо - рівноважний потенціал для іонів калію. за осі ординат - час
ка, всередині якої, як і в кіноцілій, по колу розташовуються 9 пар фібрил. Вія в одних випадках утворює розширення - зовнішній сегмент у вигляді циліндра, в інших - у вигляді колби. Відповідно перший тип фоторецептора отримав назву палички, а другий - колбочки. У колб мембрана утворює дископодібні, що накладаються один на одного випинання, дедалі менші в діаметрі в напрямку до верхнього кінця; внутрішня порожнину кожного диска повідомляється з внутрішнім середовищем колбочки. У паличок мембрани дисків повністю замкнуті і бнутреннее простір диска ізольовано від внутрішнього середовища зовнішнього сегмента палички. Диски зібрані стопкою і укладені в футляр з поверхневої мембрани зовнішнього сегмента.
В мембрани дисків обох типів фоторецепторів вбудований інтегральний фоторецептора білок - фотопігмент, концентрація якого дуже значна. Наприклад, у корів на частку фотопігмент припадає близько 80% загальної кількості білків мембрани. Все це вказує на те, що процеси фотохімічного перетворення енергії світлового стимулу відбуваються саме в дисках. Велика кількість дисків збільшує світлочутливу поверхню фоторецептора. Зорові диски утворюються протягом усього життя тварини. Вони поступово протягом декількох тижнів переміщаються по зовнішньому сегменту з наступним відділенням від нього, поглинанням і руйнуванням клітинами пігментного епітелію. Пігментний епітелій виконує ще ряд важливих функцій: перешкоджає відображенню і розсіюванню світлових променів, сприяє більш чіткому зоровому сприйняттю. Зовнішні сегменти фоторецепторів оточені відростками пігментного епітелію. При сильному освітленні частки пігменту переміщаються з епітеліальних клітин і екранують зовнішні сегменти від яскравого світла.
При реєстрації електричної реакції фоторецепторів на світловий стимул було виявлено, що палички і колбочки генерують лише гіперполярізаціонние рецепторні потенціали (див. Рис. 15.17, Г). Вимірювання провідності до освітлення і під час дії світлового стимулу показало, що в темряві мембрана зовнішнього сегмента приблизно в однаковій мірі проникна для іонів натрію і калію. Тому (див. Гл. 2) відповідно до рівняння Голдмана величина спокою фоторецепторних клітин знаходиться приблизно між рівноважним потенціалом для іонів калію і натрію. Іони натрію входять в зовнішній сегмент за своїм градієнту через канали, які в темряві відкриті.
Важливо відзначити, що при генерації темнового струму іони натрію не накопичуються у внутрішньоклітинному просторі завдяки інтенсивному функціонуванню Na+ - До+-іонного насоса. Не випадково в фоторецепторах зосереджено у внутрішньому сегменті велика кількість мітохондрій. При висвітленні натрієві канали закриваються, що призводить до зменшення темнового струму і до зрушення мембранного потенціалу в бік рівноважного калієвого потенціалу, т. Е. До гіперполяризації мембрани. Після закінчення дії світлового стимулу мембранний потенціал знову рухається в позитивному напрямку і займає рівень між рівноважними калієвих і натрієвих потенціалами. Зміни мембранного потенціалу передаються шляхом простого кабельного розповсюдження струму на базальну частину внутрішнього сегмента клітини, де модулюють процес вивільнення нейромедіатора з пресинаптичного ділянки клітини.
Розберемо механізм передачі збудження від фоторецепторного білка до іонним каналам. Згідно електрофізіологічних, біохімічних і біофізичними дослідженнями фотоактівіруе- мий білок і іонні канали в фоторецепторах просторово рознесені: внутрішньоклітинним посередником виступають молекули циклічного гуанозин-3 ', 5'-монофосфату (цГМФ). У темряві цГМФ «утримує» іонні натрієві канали у відкритому стані. Поглинання світла фотопігментом (родопсином) викликає активацію G-білка. Активоване G-білок, в свою чергу, активує фермент фосфодіестеразу, яка гідролізує цГМФ, перетворюючи його в звичайний ГМФ. Зниження концентрації цГМФ призводить до закриття натрієвих каналів і темнової струм зменшується. Після світлового стимулу в темряві під дією іншого ферменту - гуанілатциклази відбувається відновлення концентрації цГМФ. Підвищення рівня цГМФ веде до відкриття натрієвих каналів і відновленню темнового струму до початкового рівня. Те, що цГМФ є вторинним посередником в фотоелектричних процесах, дозволяє пояснити надзвичайно високу чутливість фоторецепторів. Так, поглинання рецепторной кліткою декількох фотонів, енергія яких незначна, викликає помітну електричну реакцію фоторецепторной клітини, що перевищує в десятки тисяч разів первісну енергію фотостімула. Орієнтовна, певною мірою занижена оцінка показує, що ФОТОЗБУДЖЕНОГО однієї молекули фотопігмент активує 10 молекул G-білка. Далі одна молекула G-білка активує 10 молекул фосфодіестерази, яка, в свою чергу, гидролизует 10 молекул цГМФ. Таким чином активація 1 молекули фотобелка зменшує вміст цГМФ на 1000 молекул, т. Е. Відбувається посилення реакції в 1000 разів.
Великих успіхів досягнуто у вивченні структури і фотохімічних перетворень фотопигментов в фоторецепторних клітинах при нанесенні світлового стимулу. Припущення про те, що для процесу фоторецепції потрібен якийсь пігмент, було висунуто в кінці XVI11 в. Д. Дрейпером. Незабаром після цього В. Кюне виділив чутливе до світла речовин - родопсин, яке обумовлює пурпурний колір сітківки. Їм же було показано, що після знебарвлення пігменту на світлі (пурпурний колір блідне) його колір відновлюється через деякий час в темряві за умови збереження контакту між рецепторними клітинами і пігментним епітелієм. Родопсин міститься в паличках всіх ссавців. Щільність молекул родопсину в мембрані фоторецептора вельми велика - 5 - 1012 молекул на 1 см2 мембрани. Родопсин найкраще поглинає світло з довжиною хвилі близько 500 нм (синьо-зелена область). Надалі були визначені інші пігменти.
Всі вивчені фотопігмент складаються з двох головних компонентів: білка опсина і простетичної групи, яка в одних випадках представлена ретиналь (альдегидная форма вітаміну А,), в інших - 3-гідроретіналем (альдегидная форма вітаміну А2). Дія світла на родопсин (а також на інші зорові пігменти) можна представити у вигляді такої загальної схеми:
У темряві опсин і ретиналь тісно пов'язані один з одним. Як припускають, ретиналь точно «входить» як ключ в замок в певну ділянку молекули опсина. Поглинання світлового кванта викликає ланцюг конформаційних перетворень і в ретиналь, і в опсин. Родопсин, в якому ретиналь знаходиться в формі зігнутої молекули - в формі, перетворюється в люміродопсін - нестійке з'єднання з ретиналь в / лранс-формі, з випрямленою бічним ланцюгом. Необхідно відмітити, що цис-транс-ізомеризація є єдиним ефектом, що викликається світлом в зоровому пігменті.
Усі наступні реакції відбуваються спонтанно з виділенням енергії при фізіологічних температурах (т. Е. При звичайній температурі тіла тварини). Люміродопсін перетворюється в метародопсін. Змінена форма опсина, очевидно, викликає активацію G-білка, що запускає ферментативний каскад реакцій, в результаті чого відбувається гідроліз цГМФ і закриття натрієвих каналів. Метародопсін спонтанно гідролізується до ретиналя і опсина. Транс-форма ретиналю переходить в ретинол (вітамін А |), т. е. знову йде процес ізомеризації - перетворення в цмс-форму. Після цього ДПС-ретиналь з білком опсин в темряві формують родопсин, який знову бере участь в фото- кінетичному процесі.
При дуже яскравому світлі відбувається інтенсивне знебарвлення родопсину. Для досягнення вихідного рівня родопсина потрібен якийсь час, і чутливість фоторецепторів в цей період знижена. Якщо в цей час тварина або людини помістити в темне приміщення, то все довкола мало помітні. Однак у міру відновлення рівня родопсина чутливість фоторецепторів до світла зростає і очі починають розрізняти навколишнє оточення. Цей процес пристосування називають темпової адаптацією. У нормі зміст реті- Наля в фоторецепторах в результаті фотохімічних процесів заповнюється з вітаміну А, (ретинолу), запасеного в клітинах пігментного епітелію, в які він вступає з крові. Нестача вітаміну А, в їжі призводить до уповільнення освіти ретиналю і відповідно родопсина. Це супроводжується помітним зниженням чутливості ока до світла. Око втрачає здатність до темнової адаптації, розвивається так звана «куряча сліпота».