| Головна |
| «« | ЗМІСТ | »» |
|---|
Секретообразованіе тісно пов'язане з функціонуванням секреторних клітин альвеолярного епітелію і їх структурних компонентів. Найбільш важливі ланки в ланцюзі клітинних процесів, що забезпечують освіту і виділення з клітини секреторного продукту, наступні: а) синтез різних типів РНК (рРНК, мРНК, тРНК), обумовлений генетичною інформацією, що міститься в ядрі; б) участь мРНК і тРНК в синтезі білка на рибосомах (полісомах); в) діяльність структур ЕПР в сегрегації і переміщенні секреторного продукту; г) концентрація і «упаковка» секреторного продукту структурами комплексу Гольджі.
Поняття «організація секреторного процесу» включає і сувору регламентацію секретообразованія. Різноманітні процеси синтезу і преформірованія первинних і проміжних форм секреторного продукту строго локалізовані і проходять в певні проміжки часу. Ця просторово-часова послідовність обов'язково вкладається в рамки секреторного циклу. Дуже важливою особливістю багатьох клітин є зміна їх морфологічних параметрів на всіх його етапах. Методи електронної мікроскопії та авторадіографія, а також Цитофизиологічні спостереження в ході прижиттєвого дослідження дозволили виявити конкретні етапи секреторного циклу: 1) надходження речовин - попередників секрету в тіло клітини; 2) синтез первинного секрету; 3) внутрішньоклітинний транспорт, подальша обробка і накопичення секреторного продукту; 4) видалення з клітини, виведення секрету (екструзія). Таким чином, секреторний цикл - це послідовне проходження клітиною стадій метаболічних і структурних перебудов, які забезпечують повне перетворення секреторного продукту від надходження речовин-попередників до виведення з тіла клітини готового сформованого продукту секреції.
Зміни, що відбуваються в секреторному епітелії при інтенсифікації секреції, вказують на те, що деякі процеси припадають на початок секреторного циклу - активація ядерно ного апарату збігається з інтенсифікацією транспортних процесів. На перших етапах секреторного циклу, відразу після видалення раніше синтезованого секрету, відбувається значне збільшення обсягу секреторних клітин і зміна їх форми від уплощенной до циліндричної; при одночасній інтенсифікації транспортних і синтетичних процесів змінюється біоелектрична активність клітини. Функціональне набухання обумовлено швидким надходженням в клітку осмотично активних речовин - глюкози, амінокислот і електролітів. Подовження епітеліальних клітин пов'язано з скорочувальної активністю внутрішньоклітинних цитоскелетних структур: руйнування їх за допомогою колхіцину попереджає зміна форми клітин.
На початкових стадіях секреторного циклу в кілька разів посилюється активність таких транспортних ферментів, як, наприклад, лужна фосфатаза. Зі збільшенням обсягу цитоплазми секреторною клітини збігається підвищення рівня вмісту РНК. Новоутворення РНК і транспорт її в цитоплазму викликані, в свою чергу, активацією геному клітини. Інтенсифікація транскрипционной активності хроматину проявляється в підвищенні вмісту деконденсірованного хроматину і посилення зв'язування флуоресцентних індикаторів, що вказують на дерепресія певних ділянок геному.
В ході секреторного циклу на полісомах, розташованих на поверхні каналів ЕПР, відбувається синтез пептидних ланцюгів. У полисомой на одну молекулу мРНК доводиться кілька рибосом; освіту полісомних комплексів забезпечує найбільш ефективне використання мРНК. Пройшовши через одну і ту ж серію кодонів мРНК, рибосоми синтезують однотипний білок. На рибосомах синтез білка здійснюється за 2 ... 3 хв, протягом наступних 10 хв мічені амінокислоти виявляються в складі пептидного або білкової молекули в просвіті каналів ЕПР. У клітинах молочної залози ендоплазматичнийретикулум бере участь і в синтезі молочного жиру.
Потім секреторний продукт надходить в комплекс Гольджі, де поліпептидні ланцюги формують міцели казеїну за рахунок утворення фосфатних і кальцієвих зв'язків. Структури комплексу Гольджі здійснюють ще одну функцію секреторною клітини - синтез молочного цукру (лактози). Лактоза, сформована за час проходження компонентів комплексу Гольджі, по цитоплазмі разом з гранулами казеїну переходить в порожнину альвеоли час спорожнення везикул. Очевидно, це найбільш вдалий шлях виходу молочного цукру з клітки, оскільки лактоза нездатна проникати через клітинну мембрану.
Розташовується в конденсують везикулах комплексу Гольджі секреторний продукт, що складається з казеїну і лактози, а також ліпідні глобули для виведення з клітини переміщаються в її апикальную зону. Це спрямоване переміщення пов'язане з електричної поляризацією клітин альвеолярного епітелію: внутрішньоклітинні везикули переміщуються до позитивного полюсу клітини. При дії фізіологічно активних речовин, що підсилюють виділення секрету з клітин, розвивається гиперполяризация клітинної мембрани, яка посилює ступінь поляризації клітини. Разом з цим в процесах виведення секрету за межі секреторною клітини, безсумнівно, беруть участь скоротливі елементи: цитоскелету при обробці молочної залози колхицином, речовиною, що руйнує мікротрубочки, екструзія припиняється.
Електронномікроскопічне дослідження секреторних клітин дозволяє деталізувати процес видалення секрету. Велика жирова крапля (кулька) наближається до апікальній мембрані і вбирається нею. У міру збільшення контакту з мембраною крапля все більше виступає в просвіт альвеоли. Огинаючи жирову краплю, плазматична мембрана звужується біля основи і перешнуровується, а потім відривається і потрапляє в порожнину альвеоли. Виведення з клітки білка і лактози відбувається інакше: укладені в мембрани конденсуючі вакуолі підходять до апікальній мембрані. Мембрана вакуолі з'єднується з мембраною, потім порожнину вакуолі повністю розкривається і її вміст спорожняється в просвіт альвеоли. Зауважимо, що плазматична мембрана після того, як її частина пішла на покриття молочного жирового кульки, відновлюється за рахунок розправлення на апікальній її поверхні стінки везикули, що приносить білкові секреторні гранули. Слід підкреслити, що декапитация і, отже, апокріновий тип секреції в молочній залозі не відбувається. Білок виходить з клітки по заходів- окріновому типу, а для видалення жиру використовується леммокріно- вий (лат. Lemma - оболонка) принцип екструзії.
Альвеола - структурно-функціональна одиниця молочної залози. Як показали дослідження на культивованих ізольованих клітинах молочної залози, рівень їх секреції виявляється значно нижче, ніж в клітинному співтоваристві. Тобто для збереження секреторної функції залізистих органів і, зокрема, молочної залози, необхідна цілісність клітинного комплексу як функціональної одиниці. Для більшості екзокринних залоз, включаючи молочну, такий «неподільної» одиницею є альвеола або ацинус. При аналізі принципів об'єднання клітин в альвеолярному комплексі найбільш важливими є міжклітинний взаємодія і послідовність розвитку реакцій в окремих компонентах клітинного ансамблю.
Взаємодія між секреторними клітинами обумовлено функціонуванням міжклітинних контактів, розташованих на суміжних поверхнях клітин. При внутрішньоклітинної ін'єкції флюоресцентной мітки в одну з клітин альвеоли вже через 10 ... 15 хв флуорохром виявляється в 2 ... 3 розташованих поруч клітинах, що свідчить про існування міжклітинної обміну великими молекулами. При розвитку фізіологічних реакцій альвеоли першими реагують (близько 25% клітин), і тільки потім через 10 ... 15 хв в реакцію залучаються інші клітини. Система високопроникних контактів об'єднує клітини в функціональний синцитій при збереженні клітинної відособленості. При пошкодженні сусідніх клітин залишилися цілими клітини «відключають» високопроніцаемие контакти і поширення інформації по альвеолі припиняється.
Бар'єрні властивості секреторного епітелію формуються за рахунок наявності спеціального замикательного комплексу щільного контакту, розташованого на кордоні між апікальної і латеральної зонами клітинної мембрани. Через альвеолярний бар'єр не проникають спеціальні речовини (пероксидаза і флуоресцеїну натрію). Всі речовини, що потрапляють в просвіт альвеоли, обов'язково повинні пройти через секреторну клітину.
Взаємодія між міоепітеліальние і секреторними клітинами виникає в ході функціонального сполучення процесів виведення раніше синтезованого молока і включення клітин альвеолярного епітелію в новий секреторний цикл. Виведення молока з порожнини альвеол і збудження секреторного процесу в залізистих клітинах відтворюється гормоном нейрогипофиза - окситоцином і медіатором нервової системи - ацетилхоліном. Крім того, при блокаді М-холінорецепторів реакція клітин альвеолярного епітелію не розвивається, хоча скорочення міоепітелія і виведення молока з порожнини альвеоли відбуваються. У міоепітеліальних клітинах на підставі гістохімічного дослідження виявлено можливість синтезу аце- тілхоліна, виявлена активність специфічного ферменту ацетілхолінтрансферази. Крім того, порушення синтезу аце- тілхоліна призводить до відсутності реакцій секреторного епітелію, і тільки введення його ззовні відновлює секрецію. Система інактивації ацетилхоліну представлена в молочній залозі холінестеразою, серед яких виявлені як ацетілхо- лінестераза, так і ацілгідролази. Дія антіхолінестераз- них препаратів, що попереджають гідроліз ацетилхоліну, забезпечує потенціюючий ефект. Таким чином, можна говорити про існування своєрідного механізму передачі регулюючих впливів з скорочувальної структури альвеоли (міо епітеліальної клітини) на секреторні клітини за допомогою медіатора (ацетилхоліну), який синтезується в міоепітеліі і виділяється в простір між секреторною і міоепітеліаль- ної клітинами і вловлюється холінрецептори на мембрані секреторною клітини (рис. 10.2).
Скорочення міоепітеліальних клітин розвивається при дії окситоцина і ацетилхоліну, причому клітини мають самостійними окситоциновий і холінорецептора- ми. Атропін не припиняє скорочувальної реакції альвеоли, а спільний ефект дії ацетилхоліну і окситоцину виявляється сильнішим, ніж кожного з них окремо. Істотна зміна скоротливої активності міоепітелія настає під впливом катехоламінів, які блокують скорочення альвеол у відповідь на дію окситоцину. Придушення скорочень міоепі- теліальной клітини пов'язано з реакцією 3-адреноблокатори, що запобігають подальшу скоротливу активність. Скорочення міоепітелія розвиваються при деполяризації мембрани, в тому числі і під дією електричного струму з тривалістю імпульсу не менше 350 мс.
Скорочувальний апарат міоепітеліальних клітин аналогічний такому у клітин гладкої мускулатури і представлений міо фибриллами, що заповнюють тіло і відростки клітин. Кожна міо фібрила побудована з орієнтованих в подовжньому напрямі міофіламенти товщиною від 50 до 80 нм. виявляється
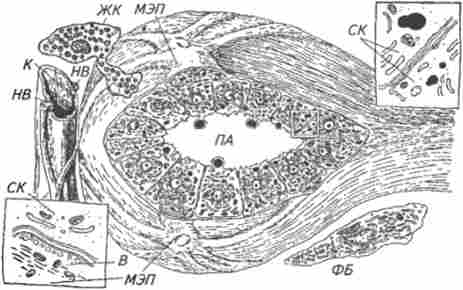
Мал. 10.2. Схема міжклітинних взаємовідносин в альвеолах молочної залози:
МЕП- міоепітеліальние клітина; СК - секреторна клітина; ПА - порожнину альвеоли; К - капіляр; В- везикули; РК жирова клітина; ФБ - фібробласт; Н В - нерви
центральна лінія прикріплення - аналог 2-мембрани. Поряд з аналогом 2-мембрани виявлені електроннощільні і електроннопрозрачние ділянки, подібні дискам А і I. Сполучені один з одним і з 2-мембраною міофіламенти заповнюють простір відростків міоепітеліальних клітин, причому в місці прикріплення до плазматичної мембрани збільшена електронна щільність хемідесмосома. Скорочення міоепітелія пов'язані з підвищенням концентрації кальцію всередині клітини. Виявлено два місця депонування іонізованого кальцію - це канали ЕПР і прімембранной везикулярне структури. При дії окситоцину і розвитку скорочення міоепітеліальние клітини іонізований кальцій виявляється в просторі між миофиламентами.
Кровопостачання альвеол істотно змінюється в ході секреторного циклу, причому розширення мікросудин і розкриття додаткових капілярів забезпечують робочу гіперемію органу. Реєстрація кровонаповнення судин, що оточують альвеоли, дозволяє виявити наявність двухфазной реакції короткочасної вазоконстрикції (звуження судин), обумовленої дією окситоцину (структурного аналога вазопресину) і тривалої вазодилатацією (розширенням судин). Інтенсивність секретообразованія тісно пов'язана з повноцінним кровопостачанням органу: об'ємний кровотік значно зростає при становленні лактації і збільшенні молочної продуктивності в лактаційний період. Порушення освіти молока при гіпогалактії зазвичай пов'язане і з відхиленнями в гемодинаміці. За даними реологічного дослідження, істотно знижується амплітуда пульсової хвилі, згладжуються ка такрота і анакрота, що вказує на спазм мікросудин молочної залози. Разом з цим падіння молочної продуктивності супроводжується зниженням шкірної температури молочної залози на 0,5 ... 1,5 ° С.