| Головна |
| «« | ЗМІСТ | »» |
|---|
Завдяки електрофізіологічних досліджень аферентних рецепторів внутрішніх органів були виявлені рецептори, що реагують на зміну хімічного складу навколишнього середовища. Серед вісцеральних хеморецепторів можна виділити наступні рецептори: змінюють свою електричну активність на зрушення pH середовища; реагують на зміну концентрації глюкози - глюкорецептори; рецептори деяких амінокислот; реагують на вміст кисню або діоксиду вуглецю: в крові - це так звані каротидні і аортальні тільця і вдихає повітрі - нейроепітеліальние тільця; сигналізують про зміну осмотичного тиску внутрішнього середовища. За винятком каротидних, аортальних і нейроепітеліальних тілець, про структуру і механізми рецепції в вісцеральних хеморецепторах відомо порівняно небагато, хоча розташовані вони в різних ділянках тіла.
Луги о- і кислото чутливі рецептори. Виявлено в слизовій оболонці шлунково-кишкового тракту тварин. Вони, мабуть, є не просто детекторами pH середовища, оскільки їх реакція залежить ще і від типу кислоти або лугу. Кислотні рецептори збільшують частоту імпульсації з підвищенням концентрації іонів водню в середовищі (т. Е. Зниженням pH), а лужні рецептори підвищують частоту аферентних потенціалів дії з ростом концентрації ОН--групи.
Глюкорецептори. Виявлено в шлунково-кишковому тракті і печінці тварин. Збільшення концентрації глюкози викликає підвищення частоти афферентной імпульсації в нервових волокнах, що утворюють ці рецептори в шлунку і тонкому кишечнику (рис. 15.20, А). Однак відомо, що зміна концентрації глюкози впливає на осмотичний тиск розчину, т. Е. Глюкорецептори можуть бути просто осморецепторами. При підвищенні осмотичності розчину за рахунок інших речовин, наприклад таких, як хлорид натрію або калію, що не виявили будь-якої реакції з боку глюкорецептори (рис. 15.20, Б, В). Порушення глюкорецептори адекватними стимулами призводило до швидкого підвищення рівня інсуліну в крові у тварин. Треба відзначити, що деякі глюкорецептори збуджують і інші вуглеводи, причому виражена специфічність реакції різних рецепторів на тип вуглеводу. Реакція глюкорецептори печінки залежить від концентрації глюкози в крові: при низьких концентраціях глюкози в аферентних волокнах реєструються потенціали дії з певною частотою, а її підвищення викликає зниження частоти імпульсації.
Амінокислотні вісцеральні рецепто- р и. Рецепторні структури, що реагують зміною імпульсної активності на амінокислоти, були виявлені в кишечнику м'ясоїдних тварин. Ці рецептори не активована іншими хімічними речовинами, а також механічним роздратуванням. Короткий латентний період відповідей на глюкозу і амінокислоти дозволяє вважати, що ці рецептори також знаходяться в слизовій оболонці внутрішніх органів тварин.
Осморецептори. У внутрішніх органах були виявлені рецептори, що реагують на зміну осмотичного тиску навколишнього їх рідини. Реакція даних рецепторів не залежала від природи речовини, за допомогою якого було викликано підвищення осмотичності розчину.

Мал. 15.20. Гістограми аффере »ггной імпульсної активності, що ілюструють специфічність глюкорецептори кишечника:
А. Реакція глюкорецептори на розчин глюкози. Б, В. Реакція на розчини NaCl і KCI (осмотичність як розчину глюкози); стрілками показано початок дії розчинів. Г. Реакція на сильне розтягнення стінки кишки; горизонтальною лінією відзначено час механічного розтягування
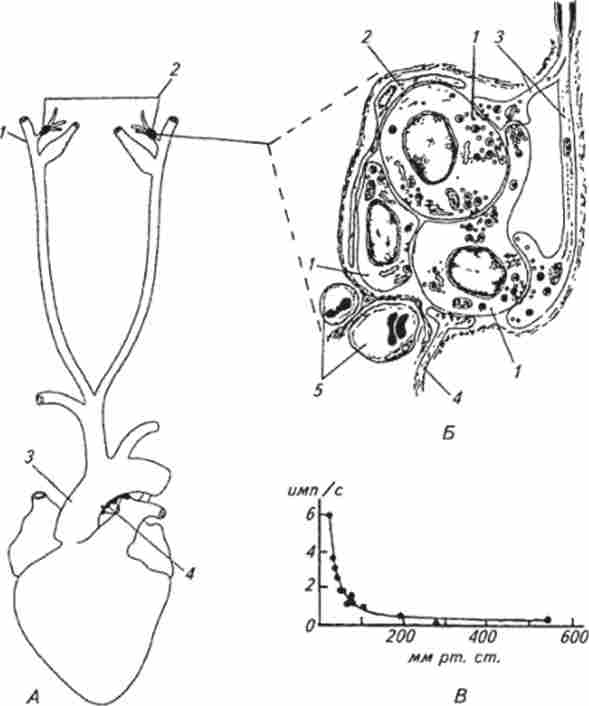
Мал. 15.21. Схема розташування каротидного і аортального тілець (Л), ділянка каротидного тільця (Б) І залежність частоти аферентних потенціалів дії каротидного тільця від вмісту кисню в крові (В):
А I - сонна артерія; 2 місце розташування каротидного тільця, 3 - аорта; 4-місце розташування аортальних тілець Б. 1 - хемореісптор- ная клітина; 2-опорна клітина; 3 - афферентное нервове волокно, 4 - еферентної нервове волокно; 5 капіляри. В. по осі ординат - частота потенціалів дії в афферентном нервовому волокні каротидного тільця, по осі абсцісс- напруга кисню в крові
Каротидного і аортальні тільця. Структура і функція цих хеморецепторів найбільш вивчені серед вісцеральних рецепторів. Вони є хеморецепторами артеріального русла і зосереджені в двох зонах: при розгалуженні загальної сонної артерії між зовнішньою і внутрішньою її гілками (Каротидні тільця) і дуги аорти (Аортальні тільця) (Рис. 15.21, /!). В основному ці рецептори чутливі до вмісту кисню в омиває їх крові, але разом з тим вони мають чітко вираженою реакцією на утримання С02 і зміна pH в кров'яному руслі. Каротидного і аортальні тільця за своєю будовою і функції мають багато спільного, однак у більшості тварин чільне місце займають каротидні тільця.
Каротидне тільце утворено сполучнотканинною капсулою, всередині якої розташовуються клітини двох типів: хеморецептори (Гломерулярні клітини) і опорні (Рис. 15.21, Б). Хеморецепторную клітини округлі, діаметр їх 10 ... 15мкм; часто вони мають цитоплазматичні відростки різної довжини. У опорних клітин також є пальцевидні відростки, які частково обплітають рецепторні клітини. Усередині капсули рецепторні клітини з прилеглими до них опорними клітинами розташовуються острівцями - кластерами, відокремленими один від одного сполучнотканинними перегородками. Співвідношення між числом рецепторних і опорних клітин всередині кластера приблизно 5: 1. Хеморецепторную клітини утворюють щільні контакти один з одним. Кровопостачання каротидних тілець здійснюється тонкими артеріями, що відходять від основної артерії. Надзвичайно густа мережа капілярів проходить через стінки капсули для бичка в кластери; всередині тілець капіляри переходять в венули. Через соединительнотканную стінку тілець проникають також аферентні і еферентні волокна, які під капсулою втрачають миелиновую оболонку, перетинають стінку кластера і утворюють різноманітні за формою аферентні і еферентні синаптичні закінчення з хеморецепторную клітинами. Таким чином, рецептором каротидного тільця є спеціальна клітка, синаптично пов'язана з першим аферентним нейроном.
У аферентних нервових волокнах каротидного тільця при нормальній напрузі кисню в крові тварини (80 ... 90 мм рт. Ст.) Реєструється спонтанна імпульсна активність. Це вказує на те, що хеморецепторную клітина спочатку деполяризована і з її пресинаптичною області відбувається викид медіатора. Мікроелектродние вимірювання підтвердили це припущення. У більшості випадків мембранний потенціал рецепторних клітин зрушать на 30 ... 40 мВ в сторону деполяризації від рівноважного калієвого потенціалу, що в основному визначає її високу чутливість до хімічного стимулу. При ступенчатообразном зменшенні напруги кисню в крові через 1 з починає збільшуватися частота аферентних потенціалів дії, яка протягом 3 з досягає максимальної величини для даного значення напруги кисню в крові. Надалі частота імпульсації залишається на незмінному рівні, т. Е. Адаптація у каротидних рецепторів відсутня. Спад напруги кисню до вихідного значення супроводжується зниженням протягом 2 ... 3 з частоти імпульсації до початкового рівня. Залежність між зменшенням напруги кисню в крові і середньою частотою аферентних потенціалів дії має вигляд гіперболи (рис. 15.21, В). Фізіологічна порогове значення напруги кисню знаходиться на рівні 75 мм рт. ст. Максимальна частота потенціалів дії в нервових волокнах каротидного тільця спостерігається при зниженні напруги кисню до 10 мм рт. ст.
При збільшенні напруги діоксиду вуглецю в крові до 100 мм рт. ст. або зменшенні pH крові до 6,9 відбувається також посилення частоти аферентних потенціалів дії в нервових волокнах каротидного тільця. Залежність між частотою і напругою діоксиду вуглецю і pH в даних межах близька до лінійної. Однак подальше збільшення цих значень супроводжується лише незначним приростом частоти аферентних потенціалів дії.
В експериментах, проведених на ізольованих хеморецеп- раторних клітинах каротидного синуса кроликів, було виявлено в мембрані клітин кілька типів іонних каналів, і в тому числі каналів, чутливих до вмісту кисню в крові. Зокрема, було показано, що при зниженні напруги кисню блокується одна з компонент іонного струму, що переноситься іонами калію. Вважають, що в мембрані хеморецепторной клітиниє калієві канали з рецепторним білком, активуються киснем. При нормальному вмісті кисню в крові білок активований і калієві канали відкриті. Зниження концентрації кисню в крові призводить до інактивації білка, закриття калієвих каналів і до зрушення мембранного потенціалу в бік деполяризації. Деполяризація мембрани викликає додатковий вихід медіатора з клітки і збудження аферентних синапсів з подальшим збільшенням частоти потенціалів дії в нервовому волокні. Питання про механізми рецепції напруги діоксиду вуглецю в крові і pH залишається менш вивченим.
Нервові волокна від каротидних тілець формують так званий синусних нерв, який представляє собою гілку язикоглоткового нерва. Тіла нервових клітин, що утворюють волокна каротидних тілець, знаходяться в шийному ганглії. Ефект стимуляції каротидних рецепторів проявляється в збільшенні дихального обсягу і подовженні фази вдиху. Імпульсація від каротидних рецепторів викликає збудження нейронів, відповідальних за фазу вдиху, і затримує його виключення.
Нейроепителиальние тельця повітроносних шляхів легенів. Аналогом каротидних і аортальних тілець в повітроносних шляхах легенів є нейроепітеліальние тільця. Вони виявлені у різних видів дихаючих повітрям хребетних тварин - від ссавців до амфібій.
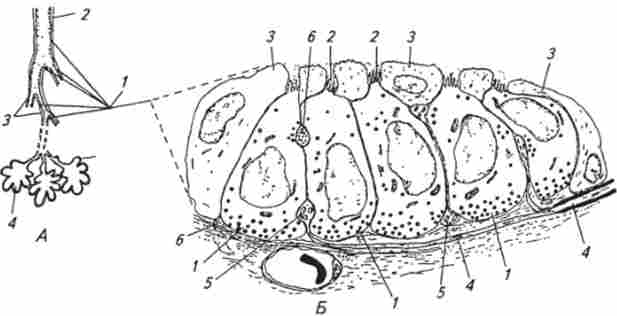
Мал. 15.22. Схема розташування нейроепітеліальних тілець (А) в повітроносних шляхах і будови нейроепітеліальние тільця (Б):
А 1 - легкі, 2-бронхи, J-бронхіола; - / - альвеола. Б. / - хемореіепторние клітини; 2 мікровілли; 3 - Клара-клітини; 4 - нервове волокно; 5 - афферентное нервове закінчення; 6 - еферентної нервове закінчення
Нейроепителиальние тельця детектируют вміст кисню в порожнинах бронхів і бронхіол і розташовуються в епітеліальної вистилки слизової оболонки їх стінок (рис. 15.22, А).
Нейроепителиальние тельця (див. Рис. 15.22, Б) утворено групою щільно прилягають один до одного хеморецепторних клітин з явно вираженою поляризацією. На апикальном кінці клітин розташовані вії - мікровілли. Як правило, верхня частина клітин нейроепітеліальних тілець у різних тварин покрита шаром сплощені Клара-клітин так, що мікровілли рецепторних клітин повідомляються з повітряною порожниною через заповнені слизом вузькі пори між Клара-клітинами. У базальної частини хеморецепторних клітин міститься велика кількість гранульованих везикул. Іннервація нейроепітеліальних тілець здійснюється нервовими волокнами, що входять до складу блукаючого нерва. Тіла нервових клітин, що утворюють ці волокна, знаходяться в вузлуваті ганглії. Нервові волокна формують на клітинах нейроепітеліальних тілець аферентні і еферентні синапси. Причому часто від аферентного аксона, що утворює аферентних синапс, відходить колатералью, що закінчується еферентних синапсом. До базальних областям нейроепітеліальних тілець підходять капіляри. Так само як у хемочувствітельності клітин каротидних тілець, в мембрані клітин нейроепітеліальних тілець були виявлені калієві іонні канали, що реагують на зміну концентрації кисню в навколишньому середовищі. Нестача кисню викликає закриття калієвих іонних каналів хемочувствітельності клітин і відповідно деполяризацию їх мембран. Деполяризація, в свою чергу, посилює викид медіатора з клітин з наступним порушенням аферентних синапсів і генерації в відходять від них нервових волокнах серії потенціалів дії. Треба відзначити, що крім участі в рефлекторної регуляції функціонування легенів через центральні нервові структури нейроепітеліальние тільця можуть регулювати роботу легеневих структур. Виявилося, що хеморецепторную клітини, що утворюють нейроепітеліальние тільця, одночасно є нейроендокринними клітинами. Вони можуть здійснювати синтез і секрецію таких біологічно активних речовин, як серотонін, кальцитонін-ліберинів, бомбезин, гастрин-ліберинів і ін. Виділяючи з клітин під впливом різних факторів, що знаходяться у вдихуваному повітрі (в тому числі і під впливом гіпоксії), в довколишні капіляри або міжклітинний простір, вони можуть викликати розслаблення або стиснення кровоносних судин і бронхів. Крім того, ряд речовин, секретіруюшіхся в клітинах нейроепітеліальних тілець, прискорюють в постнатальному періоді формування легких.