| Головна |
| «« | ЗМІСТ | »» |
|---|
Біологічні мембрани мають властиві їм характерні властивості і особливості. До найбільш важливих властивостей біомембран слід віднести замкнутість, асиметричність, динамічність, виборчий транспорт речовин через мембрану.
Замкнутість мембран. В процесі самозборки ліпідні бішару замикаються самі на себе, що призводить до усунення вільних країв, на яких гідрофобні хвости могли б стикатися з водою. Це призводить до утворення закритих відсіків в клітці (компартментов).
Асиметричність мембран. За хімічним складом зовнішня поверхня мембран відрізняється від внутрішньої. Наприклад, в мембрані еритроцитів фосфатидилхолин і сфингомиелин знаходяться у зовнішній половині бішару, а фосфатіділсерін і фосфатіділетаноламін - у внутрішній. У свою чергу, асиметрія полярних головок призводить також до асиметрії розподілу вуглеводневих хвостів, так як хвости жирних кислот фосфатіділхо- лина і сфінгоміеліна більш насичені, ніж фосфатидилетаноламін і фосфатидилсерина. Отже, плинність внутрішнього моношару буде дещо більше, ніж зовнішнього.
Найбільш асиметрично розподілені в плазматичній мембрані гліколіпіди і глікопротеїни. Вуглеводні частини гликолипидов і глікопротеїнів виходять на зовнішню поверхню, іноді утворюючи суцільне покриття на поверхні клітини - гликокаликс.
Динамічність мембран. Окремі молекули мембранних ліпідів і білків здатні вільно переміщатися в мембрані, т. Е. Вони зберігають здатність до дифузії. Так, молекули ліпідів з високою швидкістю переміщаються в площині мембрани (латеральна дифузія). Вони легко міняються місцями зі своїми сусідами в межах одного моношару приблизно 10 раз в секунду. Молекули білків, так само як і ліпідів, здатні до латеральної дифузії, проте швидкість їх дифузії в кілька разів нижче, ніж молекул ліпідів. Переміщення мембранних білків в латеральної площині може бути обмежена внаслідок тяжіння між функціонально пов'язаними білками і освіти кластерів, що в кінцевому рахунку призводить до їх мозаїчній розподілу в ліпідному шарі.
Крім цього, молекули білків і ліпідів дуже швидко обертаються навколо своїх поздовжніх осей (обертальна дифузія). Перескок ліпідних молекул з одного моношару в інший (шльопанці) здійснюється рідко, а білки, мабуть, до такого перескоку взагалі не здатні. Причина виключно повільного фліп-флопа полягає в його енергетичної невигідності, оскільки необхідно перенести полярну головку молекули ліпіду через гідрофобну область бішару. Рухливість ліпідних молекул тісно пов'язана з фазовими переходами в мембрані, т. е. зміною її стану з рідкокристалічного в кристалічний (або гелевидний). Основним чинником, що викликає фазові переходи мембранних ліпідів, є зміна температури середовища. Значення температури, при якій відбувається перехід даного липида з кристалічного в жидкокристалличне стан (і назад), називається температурою фазового переходу: гель - рідкий кристал (рис. 22.4).
Температура фазового переходу залежить від довжини вуглеводневих ланцюгів, наявності та положення цме-етиленової зв'язку, введення метильних груп в вуглеводневі зв'язку ланцюга ліпідних молекул. Істотно впливають на температуру фазового переходу також відмінності в будові полярних головок, а саме ступінь іонізації полярних груп, присутність у водному середовищі двовалентних катіонів (особливо Са2+).
Особливий вплив на плинність мембрани надає жорстке чотиричленне кільце холестеролу, занурене в ліпідний бішар. У еукаріотів при температурі 37 ° С холестерол обмежує плинність мембрани, а при більш низьких температурах він, навпаки, сприяє підтримці їх плинності, перешкоджаючи злипанню вуглеводневих ланцюгів.
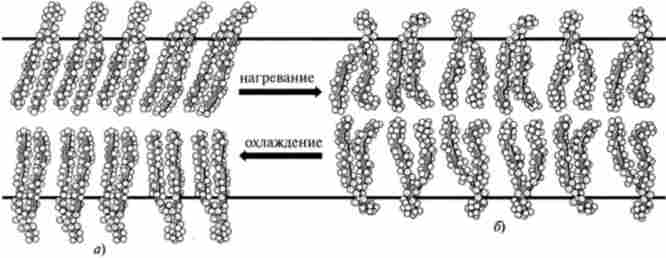
Мал. 22.4. Вплив температури на стан ліпідного бішару: а - гелеве або «кристалічна»; б - жидкокристалличне
Таким чином, температура не є єдиним чинником, що визначає фазовий стан ліпідів. Фазові зміни можуть відбуватися і при постійній температурі за рахунок зміни pH, іонного складу, присутності мембранотропної речовин, а також змін ліпідного складу бислоя. Про важливість фазового стану ліпідів для функціонування мембран свідчать широко відомі факти кореляції між температурою фазового переходу мембранних ліпідів і активністю ряду мембранно-пов'язаних ферментів.
Виборча проникність мембран. Це властивість забезпечує регуляцію транспорту в клітину необхідних молекул, а також видалення з клітини продуктів метаболізму, т. Е. Активний обмін клітини і її органел з навколишнім середовищем. Виборчий транспорт необхідний також для підтримки трансмембранного градієнта іонів, служить основою всіх біоенергетичних механізмів, визначає ефективність процесів рецепції, передачі нервового збудження і т. П.