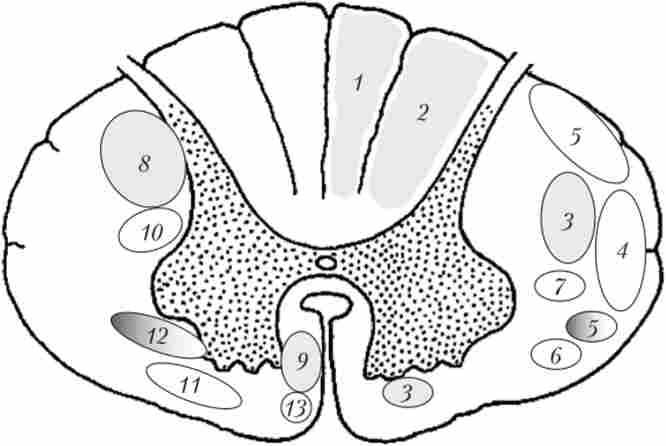
Біла речовина СМ здійснює передачу нервових імпульсів, т. Е. Забезпечує провідникову функцію. воно містить три групи провідних шляхів: Висхідні, низхідні і власні шляхи СМ (рис. 4.13).
висхідні шляху СМ передають больову, шкірну, м'язову, вісцеральний інформацію в ГМ від тулуба і кінцівок.
спадні шляху проводять керуючі команди (соматичні і вегетативні) в СМ з головного.
власні шляху з'єднують взаємодія нейронів вище-і нижележащих сегментів СМ. Це необхідно для узгодженої роботи зон сірої речовини, які управляють різними м'язами при їх одночасному скороченні (наприклад, м'язами рук і ніг під час ходьби і бігу). Крім того, в разі багатьох великих м'язів іннервують їх мотонейрони розтягнуті в ростри-каудальному напрямку на кілька сегментів. Зв'язок між ними також забезпечують власні шляхи СМ.
У задніх канатиках білої речовини йдуть висхідні шляху, в передніх канатиках - в основному спадні, а в бічних канатиках - і ті і інші. Власні шляху СМ безпосередньо примикають до сірій речовині в області як задніх, так і передніх, і бічних канатиків.
На поперечних зрізах СМ, зроблених на різних його рівнях, видно, що в верхніх сегментах білої речовини значно більше, ніж сірого; в нижніх сегментах - навпаки. Це пояснюється тим, що в грудному і особливо шийному відділах в білій речовині присутні практично всі
аксони, що зв'язують СМ з головним (як висхідні, так і спадні). Волокна ж, які досягли нижніх відділів, з'єднують з ГМ тільки поперекові, крижові і куприкова сегменти СМ. Отже, їх залишається значно менше.
Мал. 4.13.Схема розташування основних висхідних (праворуч) і низхідних (зліва) трактів білої речовини спинного мозку:
Більшість висхідних шляхів СМ організовані соматотопично (грец. Осоріо - тіло, толо (; - місце), і сигнали від різних ділянок тіла надходять в зони шкірно-м'язової чутливості головного мозку за принципом «точка в точку». Це означає, що сигнали від розташованих поруч рецепторів приходять в сусідні нейрони кори великих півкуль, і в сенсорних зонах кори завдяки цьому відбувається утворення сенсорних «карт тіла». Аналогічним чином від сусідніх ділянок рухових областей кори керуючі імпульси по спадним трактах спрямо вляются до сусідніх м'язів. Це дозволяє виявити в області моторної кори ще одну - рухову - «карту тіла».
Більшість сенсорних волокон по шляху до вищих центрів головного мозку перехрещується, завдяки чому інформація від правої половини тіла надходить в ліві сенсорні зони, а від лівої половини тіла - в праві. Перехресні чутливі волокна утворюють в СМ білу коміссуру, що лежить поруч з сірою речовиною в передніх канатиках. Нервові волокна рухових шляхів, що йдуть від головного мозку, також перехрещуються, тому права рухова область кори великих півкуль управляє рухами лівої половини тіла, і навпаки.
Як вже було сказано, на рівні СМ замикаються дуги вроджених безумовних рефлексів, що проявляються без участі інших відділів ЦНС. Однак при необхідності ГМ може регулювати їх перебіг. При цьому виділяють довільну (свідому) і мимовільну регуляцію. В останньому випадку збільшується точність рухів, а самі рухи називають автоматизованими (див. Також гл. 6). Існує також велика кількість безумовних рефлексів СМ, що запускаються вестибулярними, зоровими, слуховими та деякими іншими подразниками. Такі подразники спочатку збуджують в головному мозку сенсорні центри, імпульси від яких по спадним трактах надходять на інтернейрони і мотонейрони СМ.
У зв'язку з тим, що робота СМ протікає в постійній взаємодії з головним, поперечний розрив СМ викликає ряд важких сенсорних, рухових і вегетативних порушень. Відбувається втрата чутливості нижче області травми, так як висхідні шляху зруйновані. Спостерігається параліч м'язів, що іннервуються сегментами, що лежать каудальнее місця пошкодження, оскільки спадні тракти також не функціонують. Втрачається довільний контроль за роботою кишечника, сечового міхура і т. Д.
Характерно, що нерідко ізольований від решти ЦНС ділянку СМ відновлює здатність до здійснення безумовних рефлексів. Іноді у пацієнта можна, наприклад, викликати колінний рефлекс, хоча він не відчуває стимулу і не усвідомлює виникнення відповідної рухової реакції. Локальні пошкодження сірої речовини спинного мозку (наприклад, при пухлинах) призводять до сегментарним порушень чутливості і (або) рухових функцій на відповідному «поверсі» тіла. Найчастіше страждає робота дорсальних рогів сірої речовини шийних сегментів, що викликає погіршення і навіть втрату чутливості кистей рук.
Розглянемо конкретні найбільші висхідні і низхідні шляхи СМ.
Головні висхідні шляху наступні.
1. Спинно-бульбарні тракти проходять в задніх канатиках і називаються так тому, що з'єднують СМ з довгастим мозком (Bulbus - цибулина - застаріла назва довгастого мозку). Спинно-бульбарні тракти включають тонкий ( «ніжний») пучок, який лежить медіально, і клиновидний пучок, локалізований більш латерально (див. Рис. 4.13). Утворені ці пучки центральними відростками (аксонами) клітин спінальних гангліїв. Не входячи в сіру речовину СМ і не перехрещуючись, ці відростки піднімаються в довгастий мозок, де закінчуються на нейронах ядер тонкого і клиновидного пучків. За спіннобульбарним трактах (які називають також «спинні стовпи») максимально швидко проводиться інформація від тактильних рецепторів (при дотику, тиску), пропріорецепторов і Вісцерорецептори. Тонкий пучок проводить сигнали від нижньої частини тулуба і ніг, клиновидний - від верхньої частини тулуба і рук (в зв'язку з цим клиновидний пучок з'являється тільки на рівні грудних і шийних сегментів СМ).
2. Передній і бічний (латеральний) спинно-таламичні тракти проходять у відповідних канатиках білої речовини. Ці тракти закінчуються у верхній частині проміжного мозку - таламусе. До складу трактів входять в основному аксони вставних нейронів задніх рогів (пластини I-V) (див. Рис. 4.12), на яких утворюють синаптичні контакти відростки клітин спінальних гангліїв. Більшість аксонів интернейронов здійснюють перехрест на рівні «свого» сегмента і піднімаються до таламуса по протилежній (контролатеральной) стороні СМ. Передній спинно-таламічна тракт передає тактильні імпульси, роблячи це повільніше і в менш точної формі, ніж спинні стовпи. По суті, передній спинно-таламічна тракт є еволюційним попередником тонкого і клиновидного пучків. Латеральний спинно-таламічна тракт являє собою головний шлях для передачі больової чутливості від тулуба і кінцівок. Крім того, він проводить сигнали від температурних рецепторів. Пошкодження латерального спинно-таламичного тракту, наприклад, на правій стороні СМ призводить до втрати больової і температурної чутливості на протилежній (лівій) стороні тіла, на один сегмент нижче рівня травми.
3. Задній і передній спинно-мозочкові тракти проходять в бічних канатиках. Ці тракти також включають аксони інтеріейронов сірої речовини СМ. За цим аксонам проводиться інформація в мозочок від нропріореценторов і тактильних рецепторів. Задній спинно-мозочковою тракт не утворює перекрестов і починається від нейронів грудного ядра (ядра Кларка). Передній спинно-мозочковою тракт еволюційно більш новий; він двічі перехрещується, будучи утворений аксонами нейронів задніх рогів (в основному пластин V і VI). Інформація, яка піднімається по цим трактах, використовується мозочком для управління рухами, координації моторних актів, підтримання рівноваги і пози.
4. Спинно-ретикулярні шляхи - кілька найбільш еволюційно древніх трактів, які проводять всі види чутливості від тулуба і кінцівок до ретикулярної формації довгастого мозку і моста.
5. Спіпно-оліварний шлях проводить пропріорецептивних і тактильні сигнали в оливу (велике рухове ядро довгастого мозку). Волокна від ядер олив, в свою чергу, направляються в мозочок.
6. Спіпно-тектальіий тракт проводить різні види чутливості тіла до даху (лат. tectum) Середнього мозку. Трохи забігаючи вперед, можна сказати, що ця область головного мозку бере участь у запуску рухових реакцій на нові стимули (наприклад, поворот голови і тулуба вправо при дотику до правого плеча).
Тракти, перераховані в п. 4-6, проходять в латеральних канатиках і починаються, в першу чергу, від інтеріейронов власних ядер задніх рогів сірої речовини СМ.
Головні спадні шляху наступні.
1. Кортико-спинальний (пірамідної) тракт утворений волокнами, основна частина яких починається в руховій області кори великих півкуль (в прецентральной звивині). Найбільш товстими і швидко проводять з них є аксони гігантських пірамідних клітин п'ятого шару рухової кори. Еволюційно его наймолодший тракт СМ. Він є тільки у ссавців і добре виражений тільки в разі приматів (у людини містить близько 1 млн волокон). Пірамідний тракт проходить в вентральній області головного мозку, перехрещуючись на кордоні довгастого і СМ приблизно на 80%. Ті його волокна, які перейшли на протилежний бік, формують бічний (латеральний) пірамідний тракт. Решта аксони спускаються в СМ, де перехрещуються посегментно, утворюючи передній пірамідний тракт.
Пірамідний тракт - основний шлях для управління тонкої моторикою кистей і пальців рук, а також будь-якими довільними рухами, пов'язаними з залученням уваги і свідомості. Більшість його волокон закінчуються у власному ядрі задніх рогів, клітини якого дають аксони до проміжного ядра і мотонейронам. Отже, на шляху від кори до рухових нервових клітин зазвичай є один-три вставних нейрона. Однак у мавп і людини в пирамидном тракті присутні волокна, які контактують безпосередньо з мотонейро- нами. У людини їх 8% від загального числа аксонів; у мавп - 2%. Такі моносинаптічеськие зв'язку дозволяють здійснювати дуже швидкі і точні рухи кистей і пальців. Пошкодження пірамідного тракту порушують довільні рухи і тонку моторику пальців.
Деяка кількість волокон пірамідного тракту закінчується на нейронах рухових ядер стовбура мозку, керуючи довільними рухами м'язів голови. Відповідну частину пірамідного шляху прийнято називати кортико-нуклеарні трактом.
Решта спадні шляху, властиві СМ і пов'язані з регуляцією рухів, відносяться до екстрапірамідної системи. Екстрапіра- мідно система включає ряд підкіркових і ядерних структур, роль яких особливо велика в організації рухів, не пов'язаних із залученням уваги і свідомості. Прикладами можуть служити різноманітні рухові автоматизми, підтримання м'язового тонусу, локомоция (ходьба, біг).
2. Рубро-спінал'пий тракт бере початок від крупноклеточной частини червоного ядра (nucleus ruber) середнього мозку і далі йде в складі бічних канатиків. Волокна цього шляху закінчуються на інтернейронах задніх рогів і проміжного ядра СМ. Рубро-спинальний тракт є еволюційним попередником кортико-спинального. У людини він розвинений відносно слабко, оскільки частина його функцій приймає на себе пірамідний шлях. Рубро-спинальний тракт забезпечує збудження мотонейронів м'язів-згиначів кінцівок і гальмує мотонейрони м'язів-розгиначів. Сигнали, що передаються але волокнам тракту, також підтримують тонус м'язів-згиначів.
3. Вестибуло-спінал'ний тракт бере початок в вестибулярних ядрах довгастого мозку і моста і далі йде в складі передніх канатиків. Його волокна закінчуються на інтернейронах проміжної речовини СМ, а також безпосередньо на мотонейронах. Вестибуло-спинальний тракт пов'язаний, перш за все, з розгинанням кінцівок: він забезпечує збудження мотонейронів м'язів-розгиначів і гальмує згинання. Наявність моносінаітічних зв'язків дозволяє при втраті рівноваги робити це дуже швидко (екстрено). Крім того, сигнали, що йдуть по волокнам цього шляху, підтримують тонус м'язів-розгиначів. Особлива група ефектів вестибуло-спинального тракту - вплив на пов'язаний з підтриманням пози тонус м'язів тулуба і шиї, а також правильну ( «рівну») постановку голови і шиї.
4. Ретикуло-спінал'ние тракти йдуть від різних ядер ретикулярної формації моста і довгастого мозку. Волокна цих трактів закінчуються на інтернейронах проміжної речовини СМ. Ті, що йдуть по тракту сигнали забезпечують як збуджуючі, так і гальмівні впливи на мотонейрони вентральних рогів. Найбільший вплив вони надають на м'язи тулуба, а також регулюють роботу м'язів плечового і тазового поясів. Ретикуло-спинальні тракти еволюційно є дуже давніми і добре виражені вже у риб (управління вигинами тіла при плаванні).
5. Тектоспіпальіий тракт починається від даху середнього мозку. Функціонально він пов'язаний з уже згадуваними поворотами голови і тулуба у відповідь на нові сенсорні стимули (тактильні, зорові, слухові і т. П.).
6. Спадні вегетативні волокна являють собою велику, дифузно розподілену групу аксонів, що йде від різних структур головного мозку (гіпоталамуса, ретикулярної формації, блакитного плями і ін.) і пов'язану з емоціями, стресом, підтримкою сталості внутрішнього середовища організму. Вегетативні волокна закінчуються як на клітинах проміжного ядра, так і безпосередньо на центральних вегетативних нейронах. Імпульси, що йдуть за цими волокнам, беруть участь в регуляції роботи внутрішніх органів.
Біосинтез ліпідів, біосинтез жирних кислот - біохімія частина 2. Синтез жирів в організмі відбувається головним чином з вуглеводів, що надходять в надмірній кількості і не використовуваних для синтезу глікогену. Крім цього, в синтезі ліпідів беруть участь також і деякі амінокислоти. У порівнянні з глікогеном жири більш компактну форму зберігання енергії,
Біосинтез глікогену (глікогеногенез) - біохімія Глікоген - основна форма депонування вуглеводів у тварин - синтезується головним чином в печінці, складаючи до 6% від маси печінки, і в м'язах, де його зміст рідко перевищує 1%. Глікоген печінки виконує важливу функцію в підтримці фізіологічної концентрації глюкози в крові, перш за все в проміжках
Біосинтез ДНК і РНК - біохімія людини Синтез нуклеїнових кислот Дик і РНК з нуклеотидів відбувається в ядрі клітини і мітохондріях. Синтез ДНК носить назву реплікація, т. е. створення «дочірніх» копій- реплік, ідентичних «батьківської» ДНК. Синтез РНК носить назву транскрипція - «Переписування» інформації з матриці ДНК на матричну
Біосинтез (анаболізм) рибонуклеотидів - біохімія людини Рибонуклеотиди і дезоксірібо- нуклеотиди є мономерами ДНК, яка знаходиться в хромосомах і мітохондріях і відповідає за зберігання, передачу, трансформацію і реалізацію спадкової інформації. Практично всі живі організми, за винятком деяких видів бактерій, мають здатність синтезувати потрібні
Біологічне значення мітозу - генетика в 2 Ч. Частина 1 Мітоз лежить в основі зростання і вегетативного розмноження всіх організмів, що мають ядро, - еукаріот. Основне значення мітозу - ідентичне відтворення клітини, підтримання постійного числа хромосом, а отже, копіювання генетичної інформації. У зв'язку з цим організми, що розмножуються вегетативно,
Біологічна цінність білків злиденні - фізіологія харчування Біологічна цінність білків залежить від кількісного вмісту в них незамінних амінокислот і їх збалансованості. Розрізняють три групи харчових білків: 1) білки високої біологічної цінності - це білки, які містять в оптимальній кількості всі незамінні амінокислоти, легко перетравлюються і добре
Біохімічні основи роботи м'язів - біохімія людини Сила м'язів визначається головним чином їх розміром і здатністю повністю і координовано мобілізувати зусилля. Для успішного виконання того чи іншого виду фізичних вправ потрібна велика м'язова маса і здатність розвивати високу потужність в обмежений проміжок часу порядку декількох секунд
Білки. Структури і функції, рівні структурної організації білкових макромолекул - біохімія Білки відіграють найважливішу роль в процесах життєдіяльності. Вони є результатом експресії генів і інструментом, за допомогою якого геном управляє всіма метаболічними реакціями в клітці. Білки беруть участь в побудові клітин і тканин, здійснюють біологічний каталіз, регуляторні та скоротливі