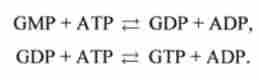
| Головна |
| «« | ЗМІСТ | »» |
|---|
Рибонуклеотиди і дезоксірібо- нуклеотиди є мономерами ДНК, яка знаходиться в хромосомах і мітохондріях і відповідає за зберігання, передачу, трансформацію і реалізацію спадкової інформації.
Практично всі живі організми, за винятком деяких видів бактерій, мають здатність синтезувати потрібні їм нуклеїнові кислоти. Синтез нуклеїнових кислот в організмі визначається швидкістю синтезу пуринових і піримідинових нуклеотидів. Синтез нуклеотидів, в свою чергу, залежить від наявності всіх складових їх компонентів - рибози, фосфорної кислоти, підстав.
Джерелом рибози і дезоксирибози є продукти перетворення глюкози в пентозофосфатному циклі. Фосфорна кислота надходить в достатній кількості з їжею.
Синтез пуринових рибонуклеотидов аденозілмонофосфата АМР (аденилат) і гуанозілмонофосфата GMP (гуанілат) починається з рибоза-5-фосфату, на якому послідовно в кілька стадій будується пуриновое ядро.
Спочатку утворюється ацикличний попередник рибонуклеотиду, а з нього після замикання ланцюга - пуриновий рибонуклеотид.
На перших двох стадіях в рибоза-5-фосфат переноситься аминогруппа від глутаміну і виходить 5-фосфо-О-рібозіламін (рис. 9.22).
Наступні стадії починаються з приєднання гліцину до 5-фосфо-Е-рибо-зіламіну і закінчуються замиканням имидазольного кільця пуринового ядра і освітою инозиновой кислоти (рис. 9.23).
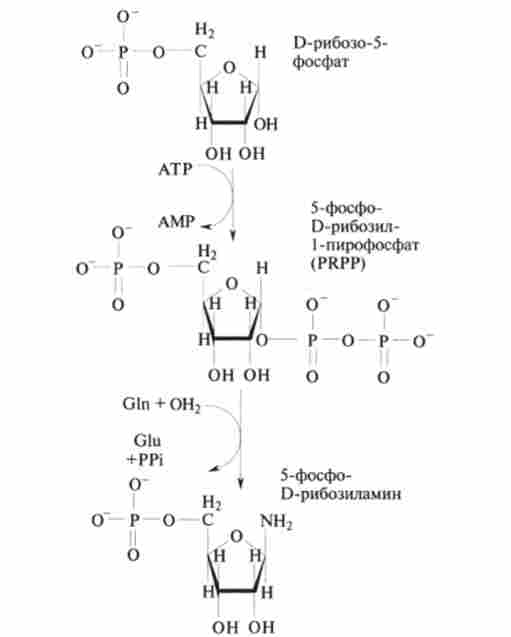
Мал. 9.22. Перетворення 0-рибоза-5-фосфату в 5-фосфо-1) -рібозіламін
Спочатку до вільної NH2-rpynne 5-фосфорібозіламін (реакція потребує доставці енергії через АТР) приєднується молекула гліцину з утворенням гліцінамідрібонуклеотіда. На наступній стадії ланцюг подовжується за рахунок приєднання формільной групи з МЛМ-метілтетрагідрофоліевой кислоти і освіти формілгліцінамідрібонуклеотіда. Потім до формільной групі останнього приєднується амидная група глутаміну і синтезується формілглі- цінамідінрібонуклеотід (реакція йде також зі споживанням енергії АТР).
На наступній стадії замикається пятичленное імідазольного кільце з утворенням 5-аміноімідазол-4-карбонової кислоти. Далі за участю аспарагінової кислоти і АТР в двухступенчатом процесі утворюються 5-аміноімідазол-4 карбоксамідрібонуклеотід і фумарова кислота.
При цих реакціях азот аспарагінової кислоти включається в 1-е положення майбутнього пуринового ядра. Останній вуглецевий атом кільця пурину приєднується до 5-NH-rpynne у вигляді формільного залишку (джерело - N-форміл- тетрагидрофолієвая кислота). Потім відбувається замикання другого кільця при відщепленні молекули води. На цьому закінчується освіту попередника пуринових нуклеотидів - инозин монофосфата IMP (инозиновой кислоти).
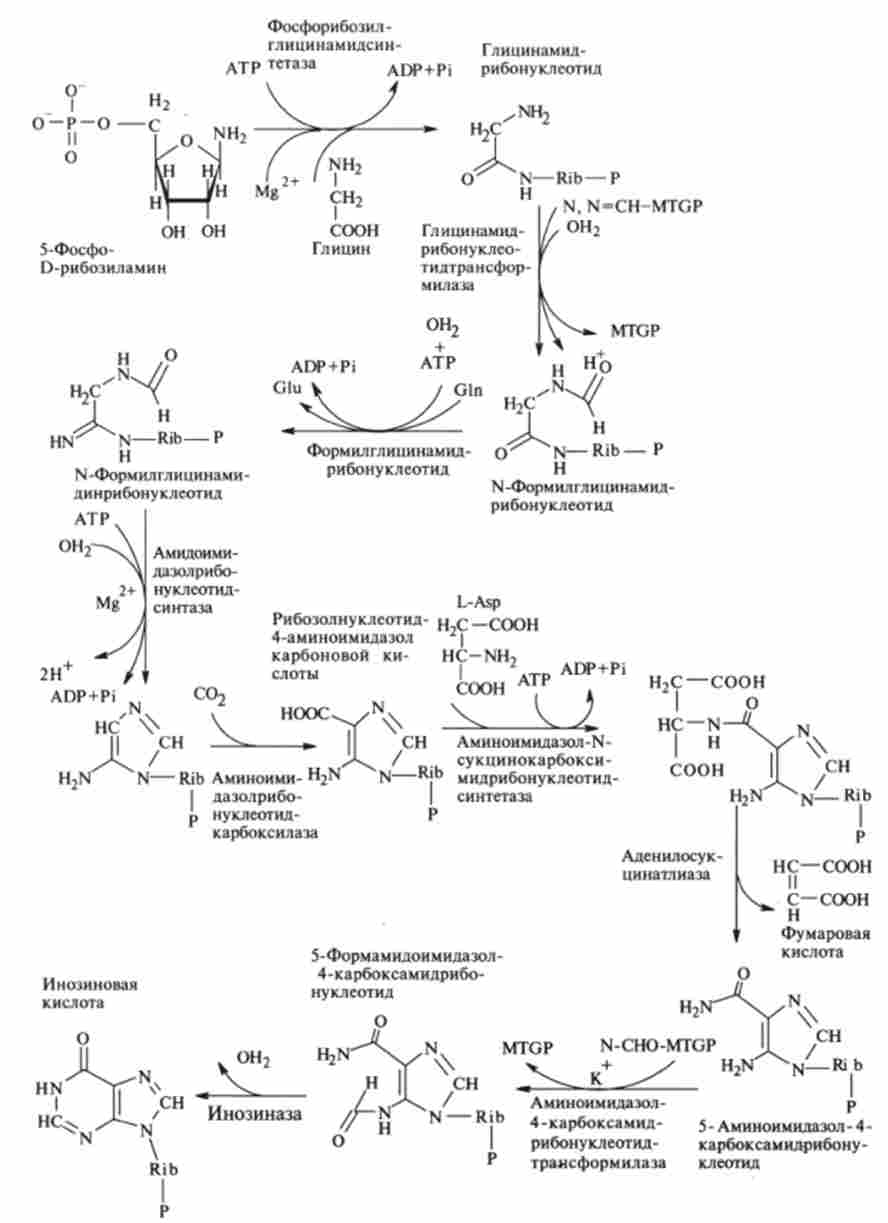
Мал. 9.23. Перетворення 5-фосфо-0-рібозіламіна в инозиновой кислоту
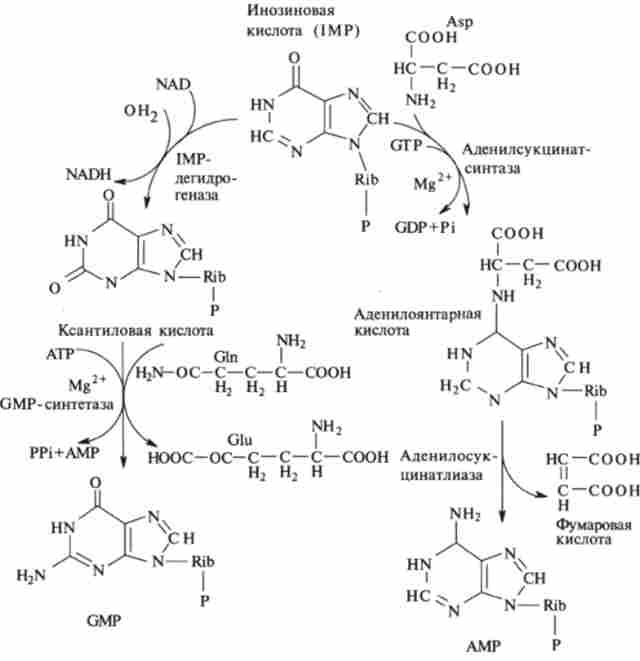
Мал. 9.24. Біосинтез пуринових рибонуклеотидов з инозиновой кислоти
Для перетворення инозиновой кислоти в аденілові (АМР) необхідне введення ще однієї аміногрупи, донором якої також є аспарагінова кислота (рис. 9.24). Джерелом енергії служить GTP. Проміжний продукт реакції - аденілоянтарная кислота.
Біосинтез GMP починається з дегідрогеназну реакції IMP, в результаті якої виходить ксантіловая кислота. В її амінірованіе використовується тільки амідний атом глутаміну. Освіта GMP каталізує IMP-дегидрогеназа і GMP-синтетаза. Освіта АМР каталізується послідовним дією аденілсукцінатсінтетази і аденілсукцінатліази.
Перетворення АМР і GMP в нуклеозідді- і нуклеозидтрифосфат протікає в дві стадії за участю нуклеозідмонофосфаткінази і нуклеозіддіфосфаткі- називаються:
Потрібно зауважити, що в клітинах існує досить тонкий механізм регуляції синтезу пуринових нуклеотидів. Їх синтез гальмується кінцевими продуктами за принципом зворотного зв'язку, т. Е. Пригніченням першої стадії перенесення аміногрупи глутаміну на PRPP. Інша особливість механізму регуляції полягає в тому, що надлишок GMP гальмує тільки свій синтез, не впливаючи на синтез АМР, і навпаки.