| Головна |
| «« | ЗМІСТ | »» |
|---|
Диференціальна експресія генів, т. Е. Регуляція їх активності в залежності від сигналів, що надходять ззовні, може відбуватися на рівні будь-якого відомого матричного процесу: реплікації, транскрипції, трансляції, а також в процесі дозрівання іРНК і поліпептидних ланцюгів, що утворюються в результаті трансляції.
Диференціальна реплікація окремих ділянок генетичного матеріалу відома у прокаріотів і еукаріотів. Індукція і подальша реплікація профага X являють собою приклад диференціальної реплікації у бактерій. ампліфікацію екстрахромосомних копій ДНК, що кодує рРНК, спостерігали в ядерцях ооцитів багатьох тварин, а також при мегаспорогенез рослин. Ампліфікація РДНК полягає в тому, що одна з її копій, що містить багаторазові повтори генів, що кодують рРНК, залишає хромосому - область ядерцевого організатора, потім багаторазово реплицируется за механізмом кільця, що котиться. Цим досягається посилений синтез рибосом в ооциті, що забезпечує ранні етапи розвитку після запліднення. Відомо, що сперматозоїд вносить в зиготу тільки ядерний матеріал і перші стадії дроблення аж до гастру- ли забезпечуються цитоплазмой, а отже, і рибосомами яйцеклітини.
У гігантських хромосомах слинних залоз двокрилих спостерігається велика політенізація окремих ділянок хромосом. Саме освіту політенних хромосом вказує на те, що реплікація в різних соматичних клітинах відбувається неоднаково. Про це ж свідчить і порівняння Ретікон - одиниць реплікації різних соматичних клітин. Розміри репліконов в ході диференціювання тканин змінюються.
Диференціальна транскрипція генів в онтогенезі добре помітна при утворенні хромосом типу лампових щіток, петлі яких, що виникають на стадії діплотени, активно транскрибуються, що добре видно на електронно-мікроскопічних препаратах ооцитів амфібій і птахів.
Інший яскравий приклад диференціальної транскрипції пов'язаний з утворенням так званих пуфів, або кілець Бальбіані, в гігантських хромосомах двокрилих. Пуфи - це характерні здуття певних дисків політенних хромосом, які утворюються в результаті локальної де- компактизации в них ДНК, що супроводжується активною транскрипцією, на що вказує інтенсивне включення 3Н-уридину в районі пуфів на препаратах політенних хромосом (рис. 10.3).
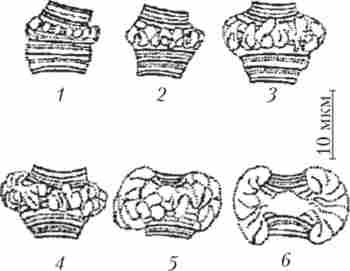
Мал. 10.3. Освіта пуфа, або кільця Бальбіані, в політенія на послідовних стадіях розвитку (1-6) личинки хирономус
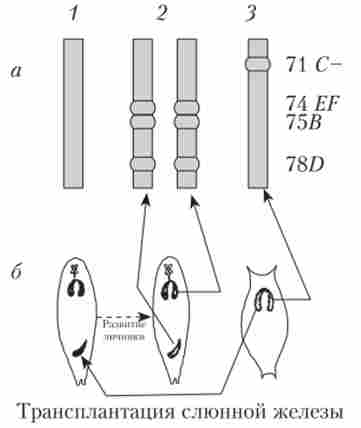
Пуфірованіе тих чи інших дисків характерно для стадії розвитку личинки. Освіта і зникнення пуфів регулює внутрішнє середовище організму відповідно до стадії розвитку. Якщо слинні залози личинки дрозофіли пересаджувати більш молодим або більш старим личинкам, то картина розподілу пуфів змінюється відповідно до тієї, яка характерна для віку реципієнта (рис. 10.4).
Мал. 11.4. Зміна характеру пуфірованія в хромосомі D. melanogaster в результаті пересадок слинних залоз більш молодим личинкам: а - гтуфи в 3-й хромосомі личинок різного віку (1-3). Праворуч - номери дисків на цитологічної карті; б - трансплантація слинної залози з більш старої личинки в молоду і порівняння характеру пуфірованія «своїх» і «чужих» хромосом при подальшому розвитку
Одним з важливих регуляторів освіти пуфів і, отже, диференціальної транскрипції генів у комах є стероїдні гормони, зокрема гормон линьки - екдизон.
Зміна структури хроматину, його декомпактізаціі, яка спостерігається при утворенні пуфів, також є одним з умов, що забезпечують диференціальну активність генів.
Диференціальна трансляція, т. Е. Синтез білка тільки на певних іРНК або регуляція синтезу білка на одній і тій же іРНК, показана для РНК-містять бактеріофагів Е. coli, а також при синтезі глобинов на стабільних іРНК без'ядерних ретикулоцитів ссавців. Надлишок геміну стимулює синтез глобина. Гемин інактивує білок, який репресує, т. е. «забороняє» синтез а- і р-ланцюгів глобіну. Деякі фракції тРНК грають роль модуляторів, задають темп трансляції; тРНК-модулятори служать лімітуючим фактором в трансляції, «впізнаючи» будь-якої унікальний кодон іРНК. Гіпотеза модулятора була запропонована на початку 1960-х рр. Г. Стент.
Можливість диференціальної трансляції полягає в існуванні стабільних іРНК, а також на збереженні іРНК в цитоплазмі у вигляді інформосом - комплексу іРНК з білками, відкритого в 1966 р А. С. Спіріним і ін.
Диференціальне дозрівання продуктів транскрипції і трансляції. Дозрівання транскриптов має на увазі модифікацію їх окремих підстав і сплайсинг про-іРНК. Кілька варіантів сплайсингу однієї і тієї ж про-іРНК досліджені для мавпячого вірусу БО 40.
Активність багатьох білків визначається їх посттрансляционной модифікацією - фосфорилюванням, ацетилюванням, а в ряді випадків розщепленням вихідної поліпептидного ланцюга на більш дрібні фрагменти.
Широко поширений механізм регуляції активності ферментів, заснований на приєднання до них молекул-ефекторів. Найчастіше в ролі ефекторів виступають кінцеві продукти ланцюгів біосинтезу, які зв'язуються з першим або з одним з перших ферментів даного метаболічного шляху і пригнічують його активність, тим самим виключаючи всю ланцюг синтезу. це інгібування кінцевим продуктом, завдяки якому регулюються відразу кілька етапів метаболізму. Кінцевий продукт зв'язується з ферментом не в його активному центрі, а в аллостерічному центрі, і така взаємодія індукує зміна (інактивацію) активного центру ферменту.
Таким чином, диференціальна активність генетичного матеріалу може забезпечуватися регуляцією різних рівнів його експресії: від реплікації до ферментативної активності білків, генних продуктів.