генотип
фенотип

індуцібельний


»



»


конститутивним
| Головна |
| «« | ЗМІСТ | »» |
|---|
Механізм регуляції транскрипції найбільш докладно досліджений у прокаріот. Ферменти клітини умовно діляться на конститутивні, присутні постійно, і адаптивні, що з'являються в результаті зміни середовища. До числа адаптивних відносяться ферменти утилізації лактози, до конститутивним - ферменти утилізації глюкози - універсального джерела вуглецю.
якщо клітини Е. coli посіяти на середовище, що містить глюкозу як джерело вуглецю, бактерії відразу ж починають його засвоювати і активно ділитися. Якщо ж помістити їх на середу з ss-галактозид - лактозою, то після деякого періоду адаптації до цього цукру бактерії почнуть його засвоювати і ділитися. За цей період відбувається індукція відразу трьох ферментів: ss-галактозидази, яка розщеплює лактозу на галактозу і глюкозу, галактозідпермеази, транспортує галактозид в клітку, і трансацетнлази, не бере участь у метаболізмі лактози. Вивчення генетичного контролю засвоєння лактози дозволило Ф. Жакобу і Ж. Моно в 1961 р сформулювати теорію оперона - основна одиниця генетичного матеріалу, регульованою на рівні транскрипції у бактерій.
лактозна оперон Е. coli (Рис. 10.5). Відповідно до теорії Ф. Жакоба і Ж. Моно ss-галактозид контролює синтез трьох згаданих ферментів, які кодують три тісно зчеплених гена: Z (Ss-галактозидазу), Y (Галак- тозідпермеазу) і А (Трансацетілазу). Найбільш важливими компонентами схеми оперон регуляції є: ген-регулятор (/), Що кодує білок-репрессор. і оператор (О), до якого має близьку спорідненість репрессор.
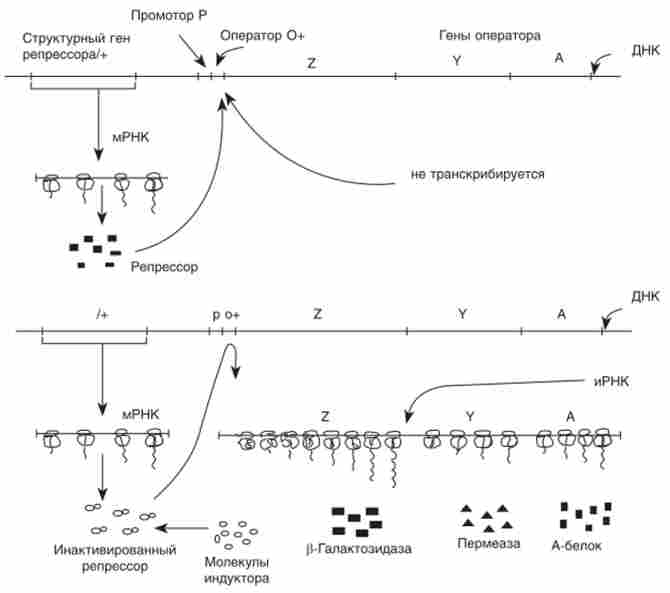
Мал. 10.5. Схема регуляції / ас-опсрона Е. coli (Індуцібельная система негативної регуляції): л - репресія: репрессор - продукт гена I зв'язується з оператором і «забороняє» транскрипцію оперона; б - індукція: молекула індуктора, з'єднуючись з репрессором, перешкоджає його зв'язування з оператором. Відбувається транскрипція оперона
Оператор і тісно зчеплені з ним структурні гени, що знаходяться під його контролем (в даному випадку 7, УА), утворюють оперон. Ген I не входить до складу оперона, хоча і може бути з ним зчеплений.
Вільний репрессор з'єднується з оператором і тим самим «забороняє» (репресує) транскрипцію оперона. Лактоза, яка в даній системі служить індуктором, взаємодіє з репрессором, в результаті чого він вже не може з'єднатися з оператором. Вільний оператор забезпечує початок транскрипції всього оперона. РНК-полімераза зв'язується з промотором і забезпечує синтез іРНК.
Таким чином здійснюється індукція оперона. Ця схема розроблена на основі вивчення мутантів з плейотропних впливом відразу на всі три ферменту, що кодуються / ас-опероном.
Гени / ас-оперону дуже рідко транскрибируются за відсутності індуктора. Індукція призводить до 1000-кратного підвищення активності всіх трьох білків. Такий же рівень дерепрессии / ас-оперону спостерігається у так званих конститутивних мутантів, які діляться на дві групи: Г - мутанти по гену-регулятору, С / - мутанти по оператору. Вони розрізняються по прояву в мерозіготах, які можна сконструювати, використовуючи Л'-Ляс-епісоми. Мутації по регулятору (Г) рецесивні (табл. 10.1). Мутації по оператору ((/) обумовлюють конститутивний прояв нормального гена 7 (І інших генів оперона), тільки будучи фізично з ним зчепленими, т. Е. В цис-положенні. Такі мутації називають цис- домінанти.
Таблиця 10.1
Фенотипи гетерозигот по мутацій (Г н 0е), що призводить до конститутивних висловом генів ? ас-оперона Е. coli (Судячи з активності ss-галактозидази)
|
генотип |
фенотип |
|
|
індуцібельний |
|
|
|
|
|
» |
|
|
|
|
|
|
|
|
» |
|
|
|
|
|
конститутивним |
Примітка. Один нз алелей гена 2 несе мутацію, інактивована тільки р-гапактозідазу.
Існування цих двох типів мутацій конститутивно добре узгоджується з припущенням про існування гена-регулятора, продуктом якого є репрессор, здатний до дифузії в клітці, і операційного ділянки, контролюючого зчитування (транскрипцію) оперону (див. Рис. 10.5).
З цією схемою (рис. 10.5) узгоджується ще один тип плейотропних мутацій - так звані полярні мутації. Якщо така мутація виникає в гені Z, ближче до кордону з О, то порушується функція не тільки гена Z, але Їй А. Якщо полярна мутація виникає в Y, то функція Z не порушена, а інак-тівіровани гени Y і А. Такий полярний ефект вказує на те, що всі три гена зчитуються як єдине ціле. Полярні мутації ідентифіковані як результат виникнення кодону-термінатора. Їх ефект пояснюється тим, що рибосоми, що транслюють загальну молекулу іРНК / ас-оперону, зустрічають кодон-термінатор, і це призводить до передчасної дисоціації транслює комплексу.
При використанні ДНК бактеріофагів X, трансдуцірующіх / ае-оперон, було показано, що репресія і індукція відбуваються саме на рівні транскрипції. клітини Е. coli в стані репресії не містять відчутних кількостей іРНК, комплементарної / ас-ДНК. Така іРНК з'являється після індукції.
Репрессор / ас-оперону, який представляє собою білок-тетрамер, виділений. Субодиниця репрессора складається з 360 амінокислотних залишків; / Ас-репрессор активно зв'язується з послідовністю в 24 п. Н. (Рис. 10.6), що має симетричну будову. Симетрія оператора дозволяє репрессора «дізнаватися» його з обох кінців, диффундируя уздовж молекули ДНК.
Молекула - тетрамер білка репрессора - дізнається оператор, наближаючись до нього з будь-якого кінця.
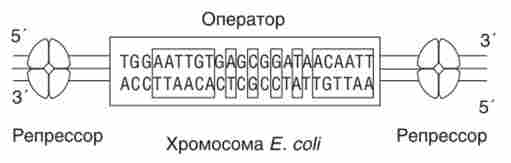
Мал. 10.6. Структура ДНК / ас-оператор, з якою пов'язується репрессор
Схема регуляції / ас-оперону, в якій низькомолекулярний ефектор лактоза служить індуктором, носить назву індуцібельной схеми негативної регуляції. Негативною ця регуляція названа тому, що білок - продукт гена-регулятора, зв'язуючись з оператором, забороняє транскрипцію т. Е. Діє негативно на експресію генів оперона.
Оперон біосинтезу амінокислот. У Е. coli і S. typhimurium весь шлях біосинтезу гистидина контролюють дев'ять тісно зчеплених генів (рис. 10.7), регульованих по оперон схемою. При індукції, яка відбувається, коли в клітці виснажується запас вільного гістидину, всі дев'ять генів Яй-оперона транскрибируются на одну молекулу іРНК розміром близько 10 000 нуклеотидів. Поява надлишку гистидина призводить до репресії Яй-оперона.
Це теж негативна регуляція, проте на відміну від / ас-оперону Яй-оперон працює по репрессібельной схемою. В даному випадку еффектор- гистидин є корепрессора. Репресія біосинтезу гистидина відбувається тільки в його присутності.
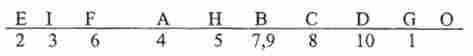
Мал. 10.7. Гени гистидинового оперона (позначені літерами), які контролюють шлях біосинтезу гистидина у S. typhimurium. Цифри - етапи біосинтезу гистидина, контрольовані даними генами
аналогічно регулюється триптофан овий оперон Е. coli, що включає п'ять структурних генів (рис. 10.8). При вивченні цього оперона був знайдений ще один спосіб регуляції - аттенюаціі (Або ослаблення). Ч. Яновський, детально дослідив трігпофановий оперон Е. coli, виявив мутанти - суперпродуцентів ферментів, що кодуються цим опероном. Мутанти несли делеции на самому початку оперона - в так званому лідерних ділянці, між оператором та ініціював кодоном першого гена trpE. При цьому втрачалася послідовність, що містить сигнал припинення транскрипції. Синтез мРНК зазвичай починається з лидерной частини, але тільки близько 10% молекул мРНК повністю копіюють йр-оперон. Передчасне припинення транскрипції забезпечує атенюатор - ту ділянку, який був втрачений у делеционного мутанггов-суперпродуцентов.
Структура лідерних ділянки іРНК йр-оперону Е. coli показана на рис. 10.8. Лідер містить ділянку зв'язування рибосоми і кодує пептид з 14 амінокислот, в якому повторюється (2 рази) залишок триптофану. Крім того, лідерних іРНК містить ділянку вторинної структури, який, мабуть, служить сигналом переривання транскрипції.
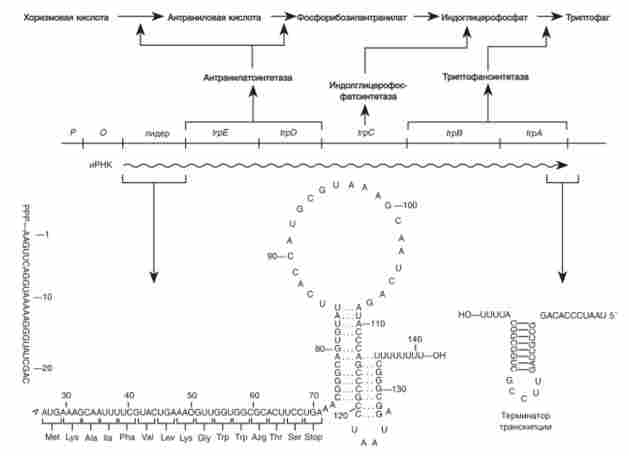
Мал. 10.8. триптофанового оперон Е. Соч
Схожі риси будови лідера знайдені в фенілаланіновой оперон Е. сон і гистидинового оперон Е. сої і 5. ? урИтіпіт. Лідер р / ге-оперону кодує пептид з 15 амінокислот, серед яких 7 залишків фенілаланіну. Лідер гистидинового оперона кодує пептид з 16 амінокислот, з них 7 - гістидин. Обидва лідери фенілаланіновой і гистидинового оперонов утворюють таку ж характерну вторинну структуру, як і лідер тріп тофанового оперона.
Мабуть, надлишок амінокислоти, синтез якої контролює оперон, призводить до накопичення відповідної аміноацил-тРНК, що забезпечує трансляцію лидерной іРНК, і рибосома, рухаючись вздовж матриці, руйнує вторинну структуру атенюатора, роблячи доступним сигнал термінації транскрипції для 6-фактора РНК-полімерази . Транскрипція припиняється. Якщо ж амінокислоти не вистачає, то трансляція лідера неможлива, і сигнал термінації транскрипції стає недоступним 6-фактору. Тоді транскрипція триває за атенюатор, і утворюється ціла молекула іРНК оперона. Таким чином, спостерігається тісний зв'язок транскрипції і трансляції в процесі регуляції оперонов біосинтезу амінокислот.
Поряд з негативною регуляцією, або репресією, відзначається позитивна регуляція. Зокрема, цей механізм продемонстрований для / аооперона Е. coli. Білок - позитивний регулятор - контролює різні системи катаболізму, до яких відноситься і / ас-оперон. Активність цього білка побічно регулює глюкоза, яка блокує транскрипцію 1ас-оперона навіть в присутності лактози. Вплив глюкози опосередковано циклічним аденозин-монофосфату цАМФ (рис. 10.9), концентрація якого в присутності глюкози знижується.
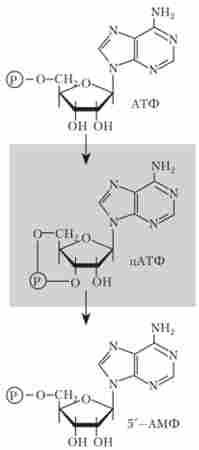
Мал. 10.9. Циклічний аденозин-монофосфат, що утворюється з АТФ і здатний перетворюватися в 5'-АМФ
Циклічний аденозин-монофосфат цАМФ зв'язується з білком - активатором катаболізму (САР - від анг. catabolite activator protein), також званим білком-рецептором цАМФ. комплекс іАМФ-САР зв'язується з промотором / ас-оперону, тим самим стимулюючи його транскрипцію.
Такий механізм позитивної регуляції вивчений для оперонов катаболізму, але невідомий для оперонов біосинтезу.