| Головна |
| «« | ЗМІСТ | »» |
|---|
еритроцити є найчисленнішими і високоспеціалізованих клітинними елементами крові. У дорослих чоловіків в 1 мкл крові міститься в середньому 5,1 млн еритроцитів, а у жінок - 4,6 млн. Основна частина еритроцитів (85%) має форму двояковогнутих дисків. Такі клітини називаються нормоцити. Їх діаметр становить 7-8 мкм, товщина в центральній області - близько 2 мкм (рис. 4.2, а). Решта 15% еритроцитів можуть мати збільшений або зменшений розмір, круглу форму (сфероціти), відростки і ін.
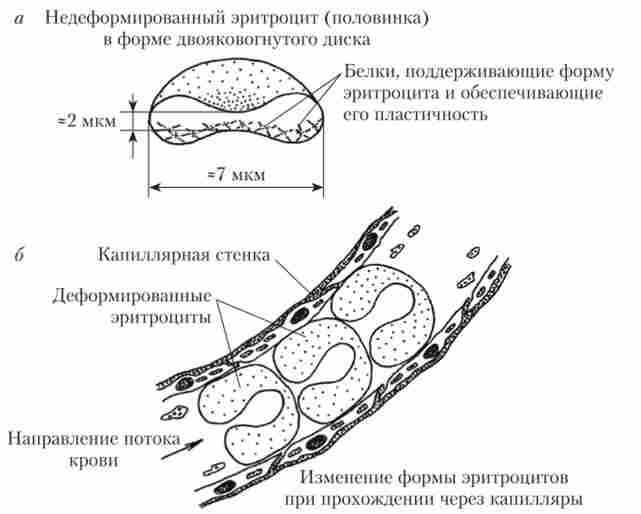
Мал. 4.2. Розміри і форма еритроцитів (А), зміна форми еритроцитів при проходженні через капіляри (Б)
Завдяки своїй будові нормоціги оптимально пристосовані до виконання газотранспортної функції. Поверхня двояковогнутого диска більше, ніж поверхня кулі такого ж обсягу, а відстань, яку повинні пройти гази від зовнішньої мембрани еритроцита до транспортних білків, - менше, ніж у клітин кулястої форми. Таким чином, дифузійна поверхня збільшується, а дифузійне відстань - зменшується. Сумарна площа поверхні всіх еритроцитів крові дорослої людини становить близько 3800 м2.
Форма еритроцитів і особливості будови мембран забезпечують їм таку властивість, як пластичність, т. е. здатність до оборотної деформації (зміни форми). Це дозволяє еритроцитів успішно проходити по капілярах, які мають значно менший діаметр (до 2-3 мкм)
і вигнуту форму (рис. 4.2, б). Пластичність еритроцитів пов'язана з присутністю в їх мембрані білків глікофорину, а в цитоплазмі - білків спектрина, які тісно взаємодіють один з одним. В процесі старіння клітин відбувається зв'язування спектрина з гемоглобіном цитоплазми. Це ускладнює нормальну взаємодію спектрина і глікофо- рина, викликає порушення форми і пластичності еритроцитів. З нор моцітов вони перетворюються в сфероціти, що є однією з причин затримки і руйнування таких клітин в селезінці. Крім цього, глико- Форін мембран еритроцитів надають їх зовнішнім новерностям гідрофільні властивості і створюють на них невеликий негативний електричний потенціал ((^ -потенціал; «дзета» -потенціал). Завдяки цьому еритроцити відштовхуються одна від одної, утворюючи в плазмі стійку суспензію.
Унікальність еритроцитів як клітин полягає в тому, що в зрілому стані вони не мають ядра і цитоплазми органел. З цієї причини еритроцити не здатні до синтезу білків або ліпідів, а також аеробного окислювального фосфорилювання. Головним джерелом енергії в зрілих еритроцитах, як і в інших клітинах, є АТФ. Ця речовина необхідно їм, зокрема для активного транспорту іонів через мембрану. АТФ в еритроцитах утворюється за рахунок анаеробного (без участі кисню) окислення глюкози (гліколізу). Це різко зменшує власні потреби червоних клітин крові в кисні, які становлять не більше 2% від усього газу, що транспортується. Іншим наслідком відсутності у зрілих еритроцитів ядра і клітинних органел є можливість розміщення додаткової кількості гемоглобіну. У зрілому еритроциті маса гемоглобіну становить близько 98% від маси всіх білків цитоплазми.
Основними функціями еритроцитів є транспортна, гомеостатична і захисна. транспортна функція полягає в здатності переносити кисень і вуглекислий газ, поживні речовини (білки і вуглеводи), біологічно активні речовини. гомеостатична функція еритроцитів обумовлена тим, що міститься в них гемоглобін може зв'язувати вуглекислий газ, знижуючи тим самим концентрацію в крові вугільної кислоти і стабілізуючи pH. Захисна функція еритроцитів полягає в їх участі в процесах згортання крові, в здатності зв'язувати і знешкоджувати деякі токсини, а також в наявності на мембранах специфічних антигенів, які роблять еритроцити учасниками ряду імунологічних реакцій.
Освіта еритроцитів (еритропоез) відбувається в кровотворних (гемопоетичних) тканинах: желточном мішку у ембріона, печінці і селезінці у плода і в червоному кістковому мозку у дитини і дорослої людини. У всіх цих органах містяться поліпотентні стовбурові гемопоетичні клітини - загальні попередники всіх клітин крові. Стовбурові клітини виробляють еритробласти - спеціалізовані клітини, з яких можуть розвиватися вже тільки еритроцити (рис. 4.3).
Еритробластів через ряд проміжних етапів росту і розвитку утворюють без'ядерні ретикулоцити, які виходять в кров і перетворюються через 24-36 год в зрілі еритроцити. Термін життя дозрілих еритроцитів - 3-4 міс., Після чого вони захоплюються і руйнуються клітинами фагоцитуючої системи селезінки, червоного кісткового мозку і печінки. Здатністю руйнувати еритроцити мають не тільки ці органи, а й більшість тканин організму, що наочно проявляється при поступовому зникненні підшкірнихкрововиливів ( «синців»). Інтенсивність еритропоезу і руйнування еритроцитів дуже велика: за одну хвилину в організмі дорослої людини утворюється 160 * 106 еритроцитів, а за добу оновлюється близько 0,8-1% від їх загальної кількості, яке становить приблизно 25 - 1012 клітин.
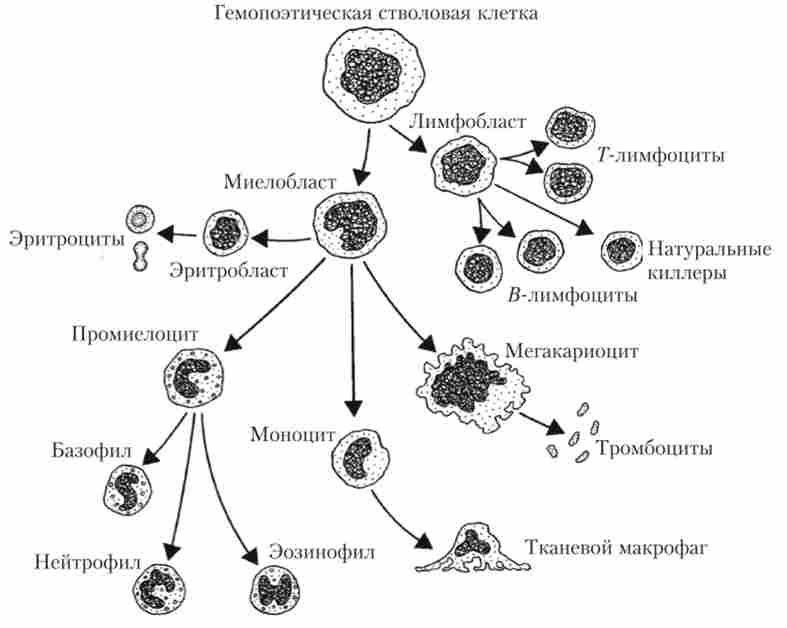
Мал. 43. Схема освіти формених елементів крові зі стовбурної гемопоетичної клітини
Еритропоез значно зростає після крововтрати, при патологічному вкороченні життя еритроцитів, а також при зменшенні кількості кисню, що надходить з повітрям (наприклад, в умовах високогір'я, коли посилений еритропоез є важливим фактором адаптації організму людини). Стимулом до прискореної диференціювання і розмноженню попередників еритроцитів в кістковому мозку є глікопротеїн еритропоетин. Головну роль в синтезі еритропоетину відіграють нирки, але в невеликих кількостях він утворюється і в печінці. Дія еритропоетину посилюється багатьма «класичними» гормонами, наприклад андрогенами, тироксином, гормоном росту. Різниця в числі еритроцитів в крові чоловіків і жінок обумовлено тим, що андрогени посилюють синтез еритропоетину, а естрогени його гальмують.
В останні роки еритропоетин здобув популярність як один із заборонених видів спортивного допінгу. Його введення в організм спортсмена в період тренувань поступово збільшує кількість еритроцитів у крові. До моменту змагань такий спортсмен отримує перевагу, краще виконуючи фізичну роботу в умовах обмеженого диханням надходження кисню. На подібному принципі заснований «кров'яний допінг». У спортсмена заздалегідь кілька разів береться кров і створюється банк еритроцитів. Перед змаганнями ці еритроцити вводяться йому в кров, що підвищує її кисневу ємкість і дає додаткову перевагу.
гемоглобін і його сполуки, сконцентровані в цитоплазмі еритроцитів, забезпечують їх життєво важливі функції - дихальну і гомеостатичну. Гемоглобін може успішно працювати, тільки перебуваючи в еритроцитах. У разі руйнування еритроцитів (гемолізу) і появи гемоглобіну в плазмі він дуже швидко зв'язується з білком гаптоглобі- ном, утворюючи комплекс, який нейтралізується клітинами фагоцитуючої системи печінки і селезінки. Період напіввиведення гемоглобіну з кровотоку становить близько 10 хв. Локалізація гемоглобіну всередині еритроцита дозволяє захистити його від впливу активних компонентів плазми, сконцентрувати і створити оптимальні умови для функціонування. Крім того, гемоглобін складним чином взаємодіє з мембраною еритроцита і елементами цитоскелету, що сприяє підвищенню його активності.
Молекула гемоглобіну складається з білкової частини (глобіну), яка включає дві пари іолііеітідних ланцюгів, і небілкової частини, що складається з чотирьох гемов (рис. 4.4).
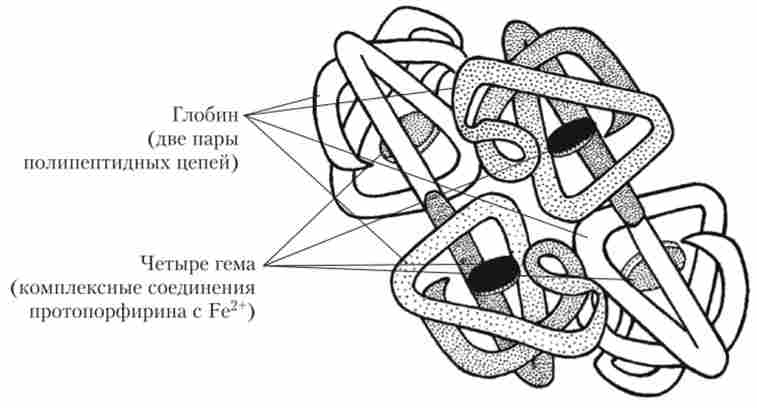
Мал. 4.4. Будова молекули гемоглобіну
Гем є комплексне з'єднання протоіорфіріна з двовалентних залізом. іон Fe2+ має здатність оборотно зв'язуватися з киснем. При цьому залізо не окислюється і залишається двовалентних. Гемоглобін, пов'язаний з киснем, називається окси- гемоглобіном. У артеріальної крові вміст оксигемоглобіну досягає 96-98% від загальної кількості гемоглобіну (останнє становить 130-170 г / л у чоловіків і 120-150 г / л у жінок).
Разом з тим, крім кисню, гемоглобін зв'язується з вуглекислим газом, утворюючи карбаміногемоглобін. Це властивість гемоглобіну забезпечує один із шляхів транспорту С02 кров'ю, а також знижує вміст вугільної кислоти в плазмі, беручи участь в механізмі підтримки pH крові на оптимальному рівні.
З'єднання гемоглобіну з киснем і вуглекислим газом можуть бути віднесені до «фізіологічним» на противагу «патологічним» з'єднанням. Прикладом патологічних сполук є карбоксігемогло- бін, який утворюється в результаті взаємодії гемоглобіну з чадним газом (СО). Карбоксигемоглобин повністю позбавлений фізіологічних функцій гемоглобіну, що призводить до тяжких отруєнь під час вступу до легені чадного газу. Іншим «патологічним» сполученням гемоглобіну є метгемоглобін, в якому залізо окислено до тривалентного стану (Fe3+). Це позбавляє метгемоглобін здатності оборотно зв'язуватися з киснем. Невелика кількість метгемогло- біна постійно присутній в крові. Перекисних сполук, вільних радикалів і інші окислювачі збільшують частку метгемоглобіну, антиоксиданти (наприклад, аскорбінова кислота) - знижують.
Важкі захворювання може викликати не «дефект» гемоглобіну, а його недостатня концентрація. Патологічні стани, при яких в крові знижується кількість гемоглобіну, об'єднані під загальною назвою «анемія» (недокрів'я). Анемія може бути пов'язана з гострою крово- іотерсй. Після такої кровонотері рідка частина крові відшкодовується протягом одних-трьох діб, а концентрація еритроцитів відновлюється до норми за три-шість тижнів. Весь цей час організм страждає від анемії.
Причиною анемії може бути також порушення функціональної активності червоного кісткового мозку, пов'язане з його радіаційним або хімічним ураженням. Таку анемію називають апластичної. Репродукція еритроцитів в червоному кістковому мозку може бути сильно уповільнена при нестачі вітаміну В12 або фолієвої кислоти, в цьому випадку формуються занадто великі червоні клітини крові різноманітної форми (мегалобластов). Збільшення розмірів не компенсує уповільнення репродукції еритроцитів. Цей вид анемії називається мегалобластной анемією.
Анемію може викликати синтез «дефектного» ^ -гемоглобіна, у якого пошкоджено р-ланцюжка. При низькій концентрації кисню такий гемоглобін кристалізується. Кристали подовжують еритроцит і надають йому вид серпа. Нетипова форма призводить до швидкого руйнування еритроцитів при проходженні через капіляри і розвитку анемії, яка отримала назву «серповидно-клітинна». На це захворювання страждають від 0,3 до 1,0% корінних жителів Західної Африки. Зовнішній вигляд еритроцитів при різних видах анемії показаний на рис. 4.5.
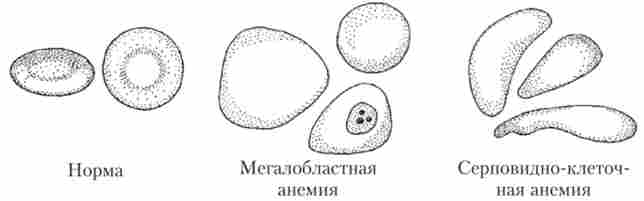
Мал. 43. Зміна форми еритроцитів при різних типах анемії